LF対談
多細胞生物の基盤、細胞接着の分子を求めて No.46(2005.9)

理化学研究所 発生・再生科学総合研究センター センター長
竹市 雅俊 氏
財団法人 千里ライフサイエンス振興財団
岡田善雄 理事長
細胞接着に関心をもったのは?
岡田 日本国際賞の受賞、おめでとうございます。竹市さんとは親しくさせてもらっているので、ちょっと話しにくいぞと思っていたんですが、えらい賞ももらわれるし、このあたりで一回ちゃんとお話を聞かせてもらわないかんということで、今回対談をお願いしました。それで竹市さんのお仕事のこと、具体的にはよく知らなかったので少し調べてみますと、細胞培養が始まった頃のことがいろいろ思い出されましてね。僕はウイルス屋だったから、培養した細胞をウイルスの宿主として使えるようになって万々歳でした。けれど、一方で多細胞生物の細胞というのは、培養してもその性格をちゃんと発揮しているという現象がいくつかあって、僕も非常に興味があった。竹市さんも細胞が接着・集合するという多細胞生物においてもっともシンプルな現象を研究されはじめたわけでしょ。これ最初ね、今みたいに展開すると思っていた?
竹市 思っていませんでしたね。要するに目の前で起こっていることを解くというだけの話で…。
岡田 それが結局、胚の組織化というか、発生における一番基本的なところにも大きく関与しているという話になったから、最初のきっかけはどうだったのかなと思いましてね。
竹市 それは偶然というか…。最初、僕が京大の岡田節人先生の研究室に入ったときは眼の水晶体の分化をやろうと思っていました。でも、その頃、岡田さんはすでに細胞接着のことをやっていたんです。腎臓の組織をバラバラにしても再構築しますが、それはもともとの細胞によるものなのか、新しい細胞によるものなのかわからないですよね。それを特定の細胞を抗体で染めて、もともとの細胞が再構築したことを初めて証明していた。細胞接着研究のカルチャーがすでに岡田研究室にあったんです。
岡田 竹市さんが接着のことをやろうと思ったのはどうして?
竹市 きっかけは単純で…。水晶体は前と後ろで細胞が違うんですね。前が水晶体上皮で、後ろが水晶体線維。ところが、手術的にひっくり返すと前の細胞だったものが後ろの細胞になる。網膜の側を向くと変わるので、これは網膜から何か因子が出ているに違いない。その因子を探そうと思ったんです。それで、網膜を培養してその培養液に水晶体の細胞を入れた。何か起こるはずでしょ。
岡田 そういうことをやっていたわけ?
竹市 でも、何も起こらない。結局、その研究はうまく実らなかったけれど、網膜の培養液を入れるのと入れないのとで、細胞の培養皿に対するくっつき方が違うことに気づいた。それがきっかけです。培養液を入れると接着が遅く、何もないと一瞬でくっつく。昔から細胞の接着にはカルシウムイオンやマグネシウムイオンがいるとされていたけれど、そんなものなしでも一瞬でくっつく。培養液を入れると、初めてカルシウムとかマグネシウムがいると。何やらおかしな話ですよね。でも、答えは簡単でした。培養液には細胞がいろんなタンパク質を分泌しているでしょ。そのタンパク質がまず培養皿に吸着する。その上に細胞がくっつくんです。結局、タンパク質が吸着した培養皿に細胞がくっつくにはマグネシウムイオンがいるとわかった。タンパク質がなかったら何もいらない。非生理的な現象ですね。それが僕の最初の論文で、さらにカルシウムイオンは細胞がお互いにくっつくのに必要らしいとおぼろげにわかってきたんです。
岡田 そのときは、まだ助手のとき?
竹市 助手のときです。細胞が培養皿にくっつくのとお互いにくっつくのでは違うシステムが働いていると。
岡田 1つの結論はついたわけですね。
竹市 自分でも新しいことを見つけたと思ったけれど、それ以上なかなか研究は進まない。接着にはどういうタンパク質の分子が必要かとか。要するに、もう壁です。そんなとき、岡田節人さんからそろそろ留学だなという話があって、1974年に米国のカーネギー研究所に行く。そこから、今の研究が始まります。結局、細胞接着に関心をもったのは、まず目で見ていて顕微鏡の下で何かおかしなことが起きていることに気づいたのと、研究室のカルチャーにそういう問題に関心を引き起こさせる何かがあったという2つのファクターによると思いますね。
細胞接着分子カドヘリンを発見
岡田 カーネギー研究所では、どんな研究をしていたの?
竹市 当時、流行っていたリポソーム(球状の脂質二重層)。細胞と細胞の接着をやっていてもなかなからちがあかない。リポソームは細胞膜にくっつくから、それを調べたら何かわかるかもしれないと。そういう研究をしているうちに、ちょっとおかしなことに気がついたんですね。細胞をトリプシン(タンパク質分解酵素)処理していったんバラバラにしても、ふつうならまたくっつきます。それがカーネギー研究所ではくっつかない。おかしいと。京大では自分でトリプシン液を作っていたけれど、カーネギー研究所ではテクニシャンが作っていました。そのレシピを見せてもらったら、僕のやらないことが1つだけあってEDTA(エチレンジアミン四酢酸)が入っていた。これはくさいと(笑)。それでEDTAを入れたり、外したり、カルシウムやマグネシウムを入れたりと、いろいろ試してみたんです。そのこと自体はリポソームの研究とは関係ない。目の前で起こっていることが不思議だから調べていると、カルシウム入りのトリプシン液では細胞をバラバラにできないんですよ。コロッと塊になるような感じで。その代わり、EDTA入りのトリプシン液だと不可逆的にバラバラになる。
岡田 それ、うれしかったでしょ。
竹市 これはいけると思った。EDTAはカルシウムを取り除く働きがある。つまり、トリプシン液にカルシウムイオンがあるかないかだけで、くっつくかくっつかないかという根本的な違いが見えた。この現象を突きつめれば絶対に重要な接着分子が見つかると確信できました。あとは本来の留学のテーマはそっちのけで(笑)。それが今の研究の始まりです。
岡田 接着分子のカドヘリンですね。それはどうやってとってきたの?
竹市 まずは細胞表面のタンパク質だけをヨード化反応で標識して電気泳動にかけました。くっつくかくっつかないかは、細胞表面に違いがあることによるはずですよね。細胞全体をつぶして電気泳動してもそんなに解像力がないから、表面タンパク質だけを比較しようと。半年くらいかかりましたが、1つだけ違いが見つかって論文も書きました。でも、それが接着分子だという証拠はなかったから、それの抗体を作って、その抗体が細胞の接着を阻害するかを見なければいけない。
岡田 そのときは何の細胞を使っていたの?
竹市 V79というチャイニーズハムスターの細胞です。それでどうするかというと、ゲーリッシュという人が細胞性粘菌の接着分子を見つけたやり方ですが、あまり難しいことを考えずに、細胞全体をウサギに注射するんです。ウサギはめったやたら抗体を作る。その中に接着分子の抗体があれば、接着を阻害するはずですよね。接着を阻害したら、その抗体を吸収するタンパク質を分画する。そうすると、それは接着分子だといえます。
岡田 そういうふうにやったのか。
竹市 でも、全然抗体を作らない(笑)。
岡田 そうなの。
竹市 カーネギー研究所のときも、それから京大に帰ってきてからも作らない。抗体を作らないと、どうしようもないです。一方で、カルシウムに依存しない接着分子があることにも気がついていて、これは学生にやらせたらうまくいって論文も出した。でも、カルシウムに依存するほうは全然うまくいかない。そんなとき、パスツール研究所でマウス由来のテラトカルシノーマ(奇形癌腫)の抗体でマウスの受精卵がおかしくなるという現象が観察されていたんです。受精卵って2、4、8と割れて、8までは1個1個の割球が丸いままで見えます。それが8の後期になると割球がぴたっとくっついて境界が見えにくくなる。これはカルシウム依存の現象です。この現象をテラトカルシノーマの抗血清が阻害すると論文に書かれていて、これはカルシウムに関係するから、僕が見ているものときっと一緒に違いないと思った。
岡田 そんな話も出てきたわけや。
竹市 だから、細胞をV79からテラトカルシノーマに変えようと。まずその細胞の接着がトリプシン液にカルシウムを入れるか、EDTAを入れるかで違うかを見たら、ちゃんと違うんですね。こりゃ、どの細胞も同じだと。それで、テラトカルシノーマの細胞をウサギに注射して抗血清をとったら、期待通りに接着を阻害する。もうその抗血清の中に接着分子の抗体があるのは明らかだから、その当時、ウエスタンブロッティングの方法が始まっていたので、手製の装置を作ってやったら、その抗血清が認識しているものがわかった。それはヨード化反応で見ていたものとだいたい分子量が同じでした。もうこれだなと思って、それが抗血清の接着阻害を吸収するかを調べたんです。でも、すんなりとはいかないですよ。なんだかんだやって間接的には一応いえたので、1982年に論文を書きました。
岡田 そのときにカドヘリンという名前はつけたの?
竹市 岡田節人さんからは早く名前をつけろといわれるけど、ちょっとまだ…。要するに、その接着分子の特異抗体で見ているわけじゃないから自信がもてなかった。幸いなことに、ちょうどその頃、モノクローナル抗体技術が開発されたので、接着を阻害する特異抗体を探しました。これ、学生の卒研でやってもらったんですが、ちゃんととれた。それで、名前もいけるなと。1984年です。

竹市 雅俊 氏
理化学研究所 発生・再生科学総合研究センター センター長
1943年、愛知県生まれ。1966年名古屋大学理学部卒業後、大学院を経て、70年京都大学理学部助手に就任。74~76年米国カーネギー研究所に留学。78年京都大学理学部助教授、86年理学部教授、99年大学院生命科学研究科教授に就任。2002年より現職。専門は発生生物学・細胞生物学。細胞接着分子カドヘリンを発見。その接着機構の解明や、カドヘリン分子群の組織維持や細胞選別における役割の研究など、細胞接着の分野の世界的リーダーとして活躍する。受賞歴は上原賞、日本学士院賞、国際発生生物学会ロス・ハリソン賞、慶應医学賞、文化功労者、日本国際賞等。2000年日本学士院会員。
ファミリーとして接着分子を整理
岡田 その頃、外国でライバルはいたの?あなたのお仕事の。
竹市 エーデルマンとか、接着に関心はもっていなかったけれどパスツール研究所のジャコブのグループも。エーデルマンは真正面から細胞接着分子を研究していました。
岡田 でも結局、あなたのカドヘリンが主流となったわけでしょ。
竹市 エーデルマンはN-CAMという分子を見つけて、これはカルシウムに依存しないほうです。それからカルシウム依存性のL-CAMという分子も見つけていて、これはあとでカドヘリンの一種となりました。結局、カドヘリンという名前が定着したのは、みんなは個別的に接着分子を探していたけれど、僕はわりと全体像をつかんでいたというか…。どういうことかというと、僕がやったのはまずテラトカルシノーマを使って接着分子を同定して、特異抗体もとれた。ところが、その抗体はテラトカルシノーマの接着を阻害するけれど、他の細胞は必ずしも阻害しない。カルシウム依存性とか共通の接着機構はあっても抗原となる接着分子は違うらしい。だから、次に神経の接着分子を見つけたときにN-カドヘリンと名づけて、最初に見つけたのは上皮にあるからE-カドヘリンとした。そのようにファミリーとして接着分子を整理していったんです。この整理法がまったく正しかった。だから、僕の命名したカドヘリンが生き残ったという。
岡田 そういう全体像があなたの頭の中にあったわけですね。
竹市 なんとはなしに。しかも、面白いのは接着分子が違うと、混ぜてもくっつかない。これこそ、昔からモスコーナとかが探していた細胞選別に関わるものに違いない。それを研究室のセミナーでいっても、あまり感動はなかったようで、みんなわかっていないなと(笑)。でも、分子的な証拠は阪大の蛋白質研究所でアミノ酸情報を調べてもらって初めて…。
岡田 たくさん試料がいるといっていたでしょ。
竹市 E-カドヘリンとN-カドヘリンを一緒にもっていったんですよ。ドキドキしてね。そうしたら、アミノ酸配列に共通性があった。非常にエキサイティングで、まさにこれはファミリーだと。直感はあたったと。
岡田 そういうのを聞きたかったんや。
竹市 なんというか、サイエンティストとしての醍醐味ですよね。
脳の神経ネットワークも接着
岡田 そして90年あたりからカドヘリンと発生学的な形態形成との関係をやられましたね。受精卵から細胞が増えて、ある形づくりができる。なぜそんなふうになるのか。もう自然の芸術品の最たるもので、手も足も出んというところに風穴を開ける物質というのを出してくれたのが竹市さんでした。カドヘリンにはいくつか種類があり、ある構造との関係では必ずあるカドヘリンとの対応があってと、まあ見事な展開やったね。
竹市 そこまではできましたけどね。そのあとは、だんだん高次構造になってくるから難しい。今は神経細胞のシナプスをやっていますから、これはそうロジカルにもいかない。
岡田 神経のネットワーク構造というのが今、興味をもっているところですか。
竹市 結局、神経ネットワークだって接着ですからね。
岡田 脳に発現しているカドヘリンの種類もずいぶんたくさんあるわけ?
竹市 他のところに発現しているものは脳にもほとんど発現している。でも、シナプスの接点はほんと小さくて…。ふつうの細胞の接着だと顕微鏡で見たら、くっついているのがわかる。シナプスは、くっついているかどうかもはっきりわからない。だから、シナプスの研究というのは生理学で進んで、誰もどうやって接着しているかなんかに関心をもたなかった(笑)。でも、そのおかげでその部分が未解決のままだった。
岡田 で、まだ実験もしているの?
竹市 いや、さすがに(笑)。実験はしませんが、学生のデータは生データをできるだけ見るようにしています。そもそも顕微鏡を見るのが好きだし、自分で顕微鏡を見ないと結果も納得できない。それと人によって気づく点が違うでしょ。
岡田 ほんとにそうですね。
竹市 学生が気づくことと、僕が気づくことは違うし、それによってもっと面白いことがわかってくるかもしれない。それに、顕微鏡を覗く楽しみがなくなったらもう…。
岡田 そうか、まだまだ現役なわけですからね。今日はお忙しいところ、どうもありがとうございました。
EYES
同じタイプの細胞を結びつけ、体づくりに働くカドヘリン
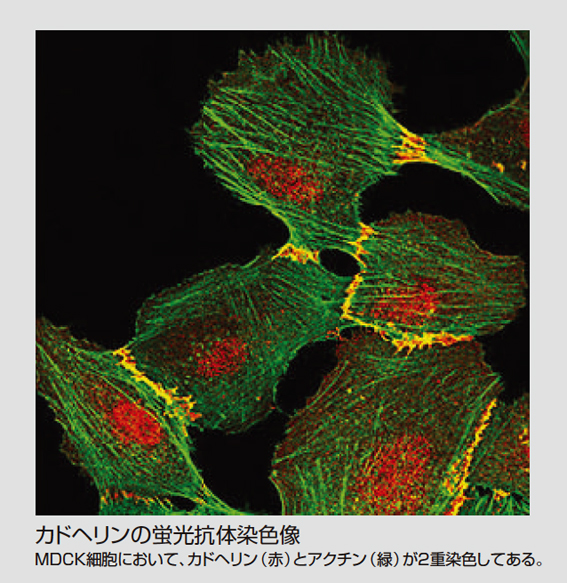
細胞同士の接着に働いているタンパク質、カドヘリンを発見
多細胞動物においては多数の細胞が集まり、接着することによって、体の組織が維持されています。皮膚や臓器など、私たちの体の組織や器官がその形を保っていられるのも、その接着のおかげです。その際、細胞同士あるいは細胞と基底膜や細胞外マトリックス(基質)との接着に働いているタンパク質を細胞接着分子と呼んでいます。
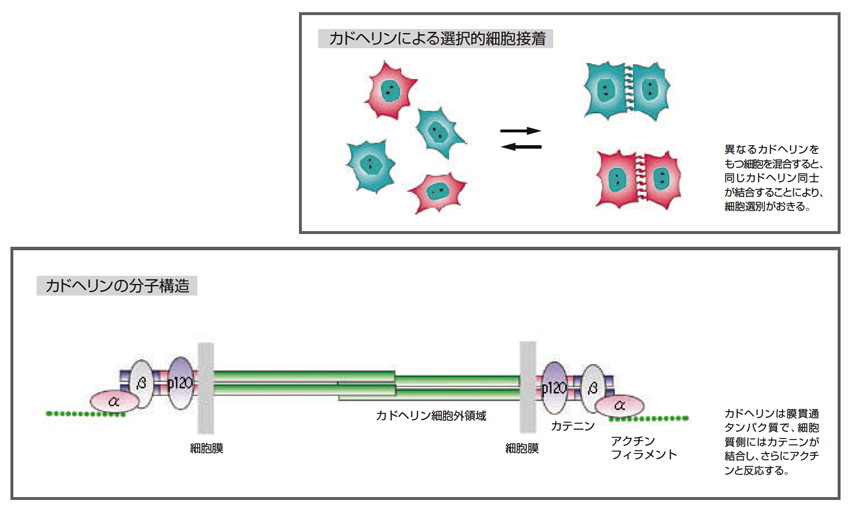
ところで、個々の組織においては、当然のように上皮細胞など同じタイプの細胞が接着しています。培養実験で組織の細胞をバラバラにして異なるタイプの細胞と混ぜても、同じタイプの細胞が再集合します。これは「細胞選別」といわれますが、その仕組みはよくわかっていませんでした。その同じタイプの細胞同士の接着に働く重要な細胞接着分子を発見されたのが、今回、LF対談にご登場いただいた竹市雅俊氏(理化学研究所発生・再生科学総合研究センターセンター長)です。
竹市氏は、カルシウムイオンに依存して働く細胞接着分子の存在を1970年代の研究を通して突きとめ、84年にカドヘリンと命名されました。さらに、カドヘリンの種類が細胞のタイプによって異なること、同じ種類のカドヘリンが選択的に結合することを解明されました。細胞選別という現象は、いろいろな細胞が発現しているカドヘリンの種類が違うことから生じていたわけです。
カドヘリンがどのように細胞同士を接着させているか、その仕組みの研究も進みました。カドヘリンは細胞膜を貫通する膜タンパク質です。細胞外の部分が同じ種類のカドヘリン同士で結合することによって細胞同士を接着させます。細胞質側の部分にはβカテニンなどの分子が結合し、βカテニンはαカテニンと結合しています。さらに、αカテニンは細胞骨格のアクチンフィラメントと反応することで、細胞の接着を安定させるといわれています。
竹市氏の研究によって、1つの受精卵から始まる個体発生の過程においても、カドヘリンが重要な役割を果たしていることがわかってきました。ある時期、ある部位において発現されるカドヘリンの種類が変化することによって、細胞はそのつど分離、再集合し、固有の働きをする組織が作られていくのです。現在、細胞接着分子として働くカドヘリンはヒトでは約20種類が見つかっています。さらに、同じ分子的特徴を有したものも含めると、その種類は120以上となり、「カドヘリン・スーパーファミリー」と呼ばれています。細胞接着分子には、他にもカルシウムイオンに依存しない「免疫グロブリンスーパーファミリー」などの分子群が見つかっていますが、同じタイプの細胞同士の接着においてはカドヘリンが主要な働きをしていると考えられています。
医学的には、がんの転移を阻止できるのではないか、という視点からカドヘリンの研究は注目されています。がん細胞にはカドヘリンの発現に異常が見られ、そのために組織から脱落し、他の臓器などに転移することがわかってきたからです。がん細胞が1か所にとどまっていれば、それを取り除くことなどによって治療は効率的に進められます。そこで、カドヘリンの発現を正常に戻す薬剤の開発が進められています。
カドヘリンの研究は、多細胞動物といっても脊椎動物が中心でしたが、竹市氏は無脊椎動物であるショウジョウバエにも対象を広げ、カドヘリンの存在を確認されています。また、現在は神経ネットワークにおけるカドヘリンの働きを研究するため、脳の海馬領域で強く発現しているカドヘリンの遺伝子を壊したノックアウトマウスなどを使った実験も進められています。カドヘリンの機能解明のさらなる進展が期待されます。