LF対談
アポトーシスはまだまだわかっていないことが多い。
できるところから攻めるしかない No.63(2011.6)

京都大学大学院医学研究科教授
長田重一 氏
公益財団法人 千里ライフサイエンス振興財団
岸本忠三 理事長
抗体だけでヒトの細胞を殺していた
岸本 僕が阪大の総長をしていた頃、10月の最初の月曜日に事務の人が「今日はノーベル賞の発表の日ですけど、資料を用意しておかないといけませんか」と聞く。「なんでや、そんなもんいらんで」と言うと、「先生と違います。長田先生です」(笑)。それが2000年ぐらい。ところが、2、3年前になると、NHKや朝日新聞が電話をかけてきて、「10月の最初の月曜日はどこにおられますか。審良(静男)先生が受賞されたら、岸本先生のお弟子さんですからぜひお話を聞きたい」と言う。世の中、変わってきたんや。アポトーシスから自然免疫、その次はどうなるかわからないですけれど。アポトーシスについては、大阪バイオサイエンス研究所の5周年記念の講演で、Fas(ファス)の抗体を打ったら肝臓の細胞が全部死んで、マウスが死んだという先生の話を聞いた。この研究に感動し、それでこの人はどうしても阪大に引き抜いてきたいと思いまして、Fasはどこから始まりましたか。先生はもともとインターフェロン、それからG-CSF(顆粒球コロニー刺激因子)というIL6もどきと誰かが言っておりましたが、それをやられて(笑)、アポトーシスになってくるわけですけれど。
長田 もともとはインターフェロン。チューリッヒ大学のワイズマン先生のところでインターフェロンのクローニング(単離)をして、その続きを日本でするとしたらそのシグナル伝達系ということになって、米原(伸)先生とインターフェロンの受容体の共同研究を始めました。そのときにどうするか。受容体に対するモノクローナル(単一)な抗体を作ろうということになった。
岸本 インターフェロンの受容体の抗体を作るために抗体を作ったわけ?
長田 米原先生、ヒトの細胞をマウスに入れて、その細胞の表面抗原に対する抗体を何千も作った。じゃあ、どうやってスクリーニング(選別)するかですけど、受容体の抗体であるならばインターフェロンが作用しなくなるはずだ。本来ならインターフェロンがその細胞に作用して、ウイルスを入れても感染しなくて細胞は死なないはずだけれどと。そうすると、インターフェロンでどれだけ処理しても細胞が死んでしまうものが出てきた。それこそ、まさにインターフェロンの受容体の抗体だと思ったわけです。だけど、コントロール実験でインターフェロンもなし、ウイルスもなしでやるとその抗体だけで殺していた。それが抗Fas抗体で、Fasというのはこの抗体を同定した米原先生が命名された。その頃は抗体がヒトの細胞を殺すと言っても、なかなか誰も信じてくれない。それならば、その抗体が認識している抗原のFasを同定しようと。私が東大の医科研から大阪バイオサイエンス研究所に移った1987年ぐらいから始めて、Fasがとれたのは91年でした。
岸本 Fasの抗体が細胞にくっつくと細胞を殺してしまう。しかも、DNAを断片化して、いわゆるアポトーシスという殺し方をする。Fasが受容体としたら、そのリガンド(結合物質)は何かということになって、それも見つかったわけですね。
長田 Fasがとれた次の年から探したけど、なかなか見つからない。何かわからない。それこそオシッコに何かないかと探したりもしました。ところが92年の12月にフランスのゴルシュタイン博士から、「ある種のキラーTリンパ球はFasを発現している細胞は殺すけど、Fasに変異のあるlprマウスの細胞は殺さない。もしかしたらFasのリガンドを出しているかもしれないけど、興味があるなら送るよ」と連絡が来たんです。すぐさま送ってもらって、リガンドを1年かけてとりました。
岸本 そこから、DNAが断片化されるまでの先生の一連の研究が進んでいくわけで、世界中でもアポトーシスに関する論文が一気に増えてきて盛んになる。アポトーシスの分野に対してノーベル賞が与えられたのは何年でしたか。
長田 2002年です。
岸本 残念ながら、先生はそこにどうしてかはいらなかったわけですけれど、それ以後、だんだんアポトーシスの分野の研究は減っていきましたよね。審良君によく言うんですけれど。自然免疫もノーベル賞が出たら終わりやなと(笑)。
長田 そうですね。それまでは国際会議に行っても、いつも人があふれんばかりだったのが、だんだん少なくなって。だけど、わかっていないことはまだまだいっぱいある。流行を追いかける人たちなんて、そんなの気にすることないじゃないですか。
アポトーシスの研究は病気の治療につながらないか?
岸本 アポトーシスというと、木の葉が離れて落ちるとか、ある時期に胎児の指の間の細胞が死んで手ができる、というようなことを説明すると、子供でも大事なことだとわかる。生命にとっては作ることも大事やけど、消すことも大事やと。そのあたりは何がどうなっているんですか。
長田 全然わかっていない。免疫系では、増えたリンパ球がアポトーシスを起こすときとかに、FasやFasのリガンドがからんで、そこに障害が起きると自己免疫疾患だとかリンパ節が腫れあがったりするけど、根本的なところ、指ができてくるところのアポトーシスがどうなっているかも何もわかっていない。
岸本 ある時期にある細胞が死なないと体づくりはちゃんとできない。このメカニズムは個体ではまったくわかっていないんですか。
長田 患者さんがいますよ。指がちゃんとできなくて止まったような患者さんが。ものすごく不思議ですよ。なんでそこで止まるのか。
岸本 先生は、なんでそういうのをやらないんですか。
長田 簡単じゃない。今のテクニックでできるところから攻めるしかない。今はアポトーシスのシグナル伝達はこうだってところはよくわかってきて、個体の発生でも同じシグナル伝達が動いていることは間違いありませんけど。
岸本 カスパーゼというタンパク質分解酵素が活性化されるところからは一緒…。
長田 一緒なんだけど、最初にどうやってカスパーゼをオンにしているかがわからない。何がトリガー(引き金)になっているのか。ホルモンがからんでいるんじゃないかという話はありますけど。
岸本 その後に先生がやっているのはアポトーシスで死んだ細胞をどのように処理するかとか、そういうことで、僕はそれはそんなに面白くないのと違うかと、よく先生に言うんですけれど。
長田 いや面白いですよ。人が亡くなっても死体が街の中に放り出されていたら、その街は社会にならないですよね。
岸本 なんである時期が来たらアポトーシスで細胞が死ぬかとか、そういうことは面白いですよね。死んだ細胞を処理するのも大事なことやけど、それはもうだいたいわかっている。
長田 全然わかっていない。10年前に我々がやり始めたときにその分野がどれだけ混乱していたか。我々、“eat me”シグナルと言っていますけど。
岸本 死んだ細胞が貪食されるのは、死んだ細胞の上に自分を食べよという信号が出るということですよね。
長田 その信号が何なのか。10年前の論文を見ると10も20もの分子が挙げられていてもう大混乱ですよ。そのときに我々が思ったのは、ちゃんと貪食を解析できていないということで、何がどう起こっているかを1つずつ調べていこうと。
岸本 だけど、たとえばマクロファージが食べてDNAを壊すというシステムはみなあるわけで、そのDNAを壊す酵素の活性を失くしたら、マクロファージが異常になって関節炎を起こしたりするというのは実験的な遊びではないかと思うんです、そんなことはない?
長田 遺伝子のノックアウト(欠損)というのは、ある意味ですべて実験的な遊びですよ。そりゃヒトの病気ではありえない。その酵素をマウスで失くしてしまったら生まれてきませんから。それほど重要な分子なんです。だけど、そういう極端な系を作ることでわかってくることもあるわけで。
岸本 普通、対談では褒める話をするんですけど(笑)。話をFasやFasのリガンドに戻すと、それはがんの治療につながりませんか。キラーTリンパ球ががん細胞を殺すのはFas、Fasのリガンドですか。
長田 使っているものもあります。もともと白血病の細胞にはFasがいっぱい出ていて、Fasのリガンドで殺すことができる。それで白血病の治療に使えないかとスタートした。ところが…。
岸本 なんでうまくいかない?
長田 Fasは、肝臓の細胞とか普通の細胞にも出ているので、白血病の細胞だけを殺そうとすること自体に無理がある。むしろ肝臓の細胞のほうがセンシティブなぐらいで、たいへん危険だということです。
岸本 病気の治療につながるようなことはないわけですか。
長田 アポトーシスに関しては、抗がん剤に抵抗性があるものに対して。
岸本 そのアポトーシスを進めるということ?
長田 それはすごくよくなってきています。アポトーシスを抑制するBcl-2に似た分子が発現してきて、抗がん剤が効かくなくなる。それを調整してあげると…。
岸本 抗がん剤がもっと効くようになる。抗がん剤が効かなくなるのは、アポトーシスを防ぐ仕組み、がん細胞が自分を守る仕組みを強固にするからで。
長田 それは今、すごい勢いで臨床研究が行われています。そういう分子を抑えるのにどういう薬剤が開発できるか。ただ、その薬剤を使っていると、また抵抗性のあるものが出てきて追っかけっこなんですけどね。あともう1つ、病気ではアポトーシスを抑えるほうならいいだろうという研究も。それはまだこれからですけど。

長田重一 氏
京都大学大学院医学研究科教授
1949年、石川県生まれ。72年東京大学理学部卒業。77年同大学院理学系研究科博士課程修了後、同大学医科学研究所助手。77~81年スイス・チューリッヒ大学分子生物学研究所に留学。87年大阪バイオサイエンス研究所分子生物学研究部部長。95年大阪大学医学部教授、99年同大学院医学系研究科教授、02年同大学院生命機能研究科教授。07年京都大学大学院医学研究科教授。専門分野は生化学、分子生物学。FasやFasリガンド、DNase(DNA分解酵素)の解明など、アポトーシス(細胞死)の分子機構の先駆的研究で世界的に知られる。受賞は、ベーリング賞、コッホ賞、朝日賞、日本学士院賞・恩賜賞、文化功労者。日本学士院会員。
研究する場所を変えるのはいいこと
岸本 先生は2007年に阪大から京大へ移られる。みな言いましたよね。もう最後のほうなのに、なんで今頃移るんやろと(笑)。
長田 みなさんそう言われるのもわからないでもない。だけど、50代後半になってくると、自分もそうだし、研究室自体もあと何年だねと、ものすごく沈滞しません?
岸本 それは移っても同じことでは、歳は。
長田 そうだけど、場所を変えたらやっぱり違います。これまでだって、東大から大阪バイオサイエンス研究所に移ったときもエキサイティングだったし、阪大に移ったときもそうでした。動かすことによって研究室を活性化できる。京大は2年間定年を延ばしましたから、65歳までに8年です。8年っていったら、一仕事できないはずがない。全然おかしくないじゃないと。
岸本 それで京大に行ってどうですか。
長田 すごく面白くなってきています。今は死んだ細胞の自分を食べてくれという信号は、ホスファチジルセリンというリン脂質だと言われている。そのリン脂質がどうやって細胞膜の内側から外側に出てくるのか、まったくわからない。アポトーシスの過程で残されたパズルの1つなんです。それを一生懸命追いかけていると、面白い膜タンパク質がとれてきた。細胞膜に関する細胞生物学の根本の問題に乗り込んでいけそうで、それがおかしくなるといろんな病気を起こしている。今、我々が見つけてきたのは血小板でホスファチジルセリンを露出させる酵素。小児血友病の一種であるスコット症候群にもからんでいる。
岸本 まだ活発に研究は続いているわけですね。
長田 当り前です。何言っているんですか(笑)。
岸本 普通は続いていなくても当り前やと思う。先生は2000年前後に1つのピークを迎えるわけです。みんなピークを過ぎるとあとはだいたい落ちていく。ところがこの頃は、歳をとってからでも特任教授だとかいって大学にいる。僕もおるんですけど(笑)。もう辞めようと思っているけど、そういうのはどう思いますか。やっぱり人によってはいつまでもいけるものなのか。
長田 すごく無理がきいて論文を読んだってすぐ頭に入ってきたときとは、今はやっぱり違います。無理がきかなくなっています。
岸本 この頃の大学のそういうシステムは。
長田 それは人によると思います。80になってもずっと変わらない人もいますし。
岸本 無難な答えですね。若い人はどうですか。今は設備もよくなったし、研究費も十分あってと外国にもあまり出ていかない。
長田 今は私がチューリッヒに行ったときとは全然、研究環境は違う。だからといって留学は必要ないかというと、とんでもない話です。日本の考え方と向こうの考え方、サイエンスのやり方はやっぱり違います。2年でも3年でも彼らと一緒にやると全然違う。うちの人たちにはどんどん行きなさいと言っているし、けっこう行っています。だけど、日本の学生さんが外に出ようとするときに奨学金があまりないということはあります。
岸本 行かないのや、この頃は。
長田 だけど、お金をとるのも大変だし、もう1つ帰ってきてもどうなるか、今は大学のポスト自体がどんどん減っていますから。
岸本 それは年寄りが残るからで。
長田 それは言えます(笑)。それから国の方針で定員削減がありますよね。運営費交付金も10年近くにわたって毎年1%ずつ下がってきているわけですから、それはやっぱりすごく痛い。
岸本 前よりもやりにくくなっていますか。
長田 大変です。
岸本 だけど、審良君が京大に移ると言ったときは、止めとけと言えた。けれど長田先生には言えませんしね。
岸本 それもまたちょっと寂しかったんですよね。おまえは阪大の人間でもないし、勝手にやれやというのは(笑)。だけど、動くというのはいいことです。私にとってはいいことです。
岸本 そりゃいいよ。そうじゃなかったらおかしなものがたまってくる、アポトーシスをおこした細胞を掃除するシステムは必要です(笑)。研究室も移転することはいいことだと思いますか。
長田 そうだと思います。今回も、京大からこんな話があって研究室全体で移ろうと思うと言ったとき、みんな一瞬シーンとなったけど、それだったら移りましょうとなって、秘書の人から技官の人から二十数人引き連れて行きました。やっぱり場所を変えたら、少なくとも数年はすごく興奮状態ですよね。
岸本 僕はずっと大阪大学やけど、少なくともその中では変わっていますから。
長田 今のホスファチジルセリンも、京大に変わったから出てきたんだろうと。
岸本 そんなに面白いの?
長田 面白い。細胞生物学の根本で、細胞の融合にも関わっているかもしれない。エンベロープ(被膜)を持つウイルスってありますよね。あれって結局、ホスファチジルセリンがむき出しになっている。そうすると、アポトーシスの細胞とウイルスってけっこう一緒なんじゃないかと。
岸本 だけど、あまり流行になってこない。
長田 だって、我々の論文もこの間出たばかりですから。アポトーシスだってlprマウスにぶつかってワーッとなりましたけど、それまではパッとしなかった。ですから、もうちょっと待ってください。
岸本 今日はどうもありがとうございました。
EYES
細胞に備えられた死のシステム、アポトーシスの分子機構の解明に貢献
自己免疫疾患にも関わるFas、Fasリガンドの発見、
DNA断片化のメカニズムの解析などを進める
私たちの体は約60兆個の細胞で構成されていますが、細胞の増殖や分化と同様に、細胞の死もまた生体のホメオスタシス(恒常性)の維持にとってなくてはならないものです。皮膚細胞は約1カ月ごとに古い細胞が死んで、新しい細胞に置き換わります。肝臓の細胞も1年ですべて新しくなっているといいます。こうした生体の能動的な細胞死をアポトーシスと呼んでいます。1972年にイギリスの病理学者カーらによって名付けられました。
アポトーシスという言葉は、「離れて、落ちる」というギリシャ語に由来します。細胞が膨張し、溶解する病的な死、ネクローシス(壊死)とは異なり、細胞の縮小、細胞やDNAの断片化などの形態的な特徴が見られます。アポトーシスは、プログラム細胞死とも呼ばれます。オタマジャクシの尻尾がなくなるなど、多細胞生物の発生、成長の過程では、特定の細胞が特定の時期に死ぬことによって体づくりは行われます。発生学では、そうした死をプログラム細胞死と呼んでいました。現在では、アポトーシスとプログラム細胞死は、細胞に備えられた死のシステムによる細胞死として区別せずに使われるようになりました。
2002年、アメリカのホロヴィッツらが「器官発生とプログラム細胞死の遺伝制御に関する発見」でノーベル医学・生理学賞を受賞します。体長1㎜ほどの線虫の発生、成長における細胞死のメカニズムの研究が評価されました。その受賞の前の90年代は、アポトーシスの研究が一気に盛んになった時期でした。その一翼を担ったのが、今回、LF対談にご登場いただいた長田重一氏(京都大学大学院医学研究科教授)です。長田氏は、主に免疫系においてアポトーシスを誘導するFas、Fasリガンドの発見や、DNA断片化のメカニズムの解析などによって、アポトーシスの分子機構の研究発展に多大な貢献をされました。
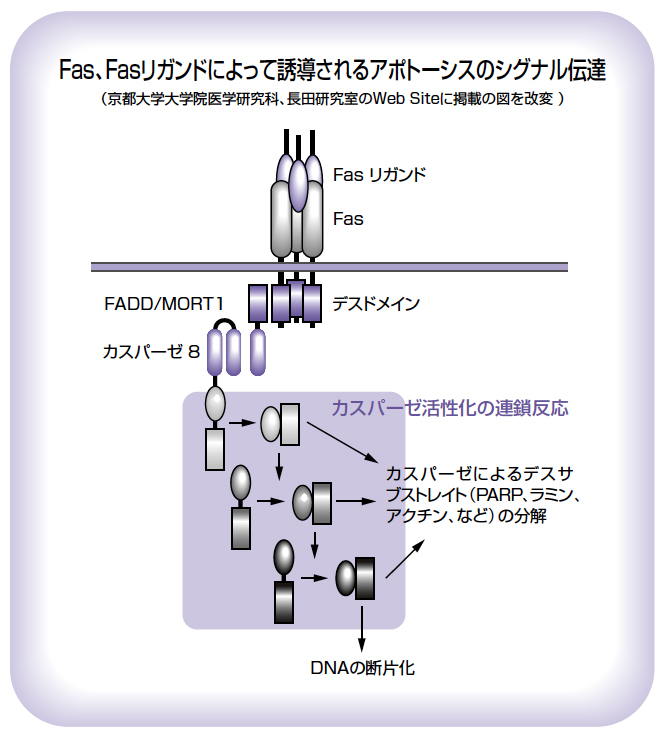
長田氏は、東京大学医科学研究所の助手だったとき、東京都臨床医学総合研究所にいた米原伸氏とインターフェロン受容体の共同研究をはじめました。米原氏は、偶然、細胞に死をもたらす抗体を作られ、その抗体が認識する抗原タンパク質をFasと名付けられました。そして、長田氏は大阪バイオサイエンス研究所に移った後の1991年にFasの遺伝子を単離されますが、それはTNF(腫瘍壊死因子)の受容体によく似たものでした。また、Fasを発現している細胞が抗体の刺激によって死ぬとき、DNAの断片化が起こることも発見され、Fasはアポトーシスを誘導する受容体であると論文で発表されました。
長田氏は、異常なTリンパ球が蓄積してリンパ節や脾臓が腫れあがるなど、自己免疫疾患のモデル動物とされる自然発生のlprマウスにFas遺伝子の変異があることも突き止められ、自己免疫疾患との関係からFasは世界的に注目されるようになりました。1993年にはFasに対するリガンド(結合物質)も発見されます。フランスの研究者の協力を得て、キラーTリンパ球が発現している、TNFに似た膜タンパク質であることを報告されました。キラーTリンパ球は、ウイルスに感染した細胞やがん細胞などの排除に働きます。免疫系のホメオスタシスの維持や、キラーTリンパ球の攻撃におけるFas、Fasリガンドの役割が次第に明らかにされていきました。
Fasを介して細胞内にどのようにアポトーシスのシグナルが伝達されるか、その分子機構の解明も進みました。FasリガンドがFasに結合すると、カスパーゼ8と呼ばれるタンパク質分解酵素が活性化され、さらに下流のカスパーゼ3などの活性化が引き起こされて、DNAの断片化などアポトーシスに特徴的な変化が起こります。長田氏は、大阪大学に移ってからはDNA断片化の解析にも取り組み、CADというDNA分解酵素と複合体を作っている阻害因子(ICAD)がカスパーゼによって切断されることで、CADが活性化されることを突き止められました。その論文は、1998年に発表された論文の中で引用回数が世界一にもなりました。その後は、アポトーシスを起こした細胞が免疫細胞のマクロファージにどのように貪食されるか、などの研究に取り組まれています。
アポトーシスは、F a sリガンドの他にもTNFやホルモン、ストレスなどさまざまな外的、内的な要因によって引き起こされます。また、増殖や成長に関わる因子が止まることによっても起こります。アポトーシスのさらなる研究発展、その医療応用への展開が期待されます。