LF対談
私の最大の興味はタンパク質の寿命で、
タンパク代謝を研究するならこれだと No.65(2012.2)

東京都医学総合研究所所長
田中啓二 氏
公益財団法人 千里ライフサイエンス振興財団
岸本忠三 理事長
細胞内でタンパク質を分解する巨大な酵素を発見
岸本 20世紀の分子生物学というのは、タンパク質がどう作られるか、あるいはそれがどういう働きをしているか、という研究が主でした。田中先生はそれを生のほうのタンパク質の研究やと言われていますね。それが21世紀に入って、タンパク質はどう壊されるかというのが非常に重要やとなってきました。その代表が先生のユビキチン・プロテアソーム系と、大隅良典先生のオートファジーですけど、それはそうですな、作るばっかりで壊されなかったらどうにもならないわけで。先生がプロテアソームの研究を始められたのはいつ頃からですか。
田中 私の先生は市原明先生でアミノ酸代謝を研究されていました。私が1972年に徳島大学の大学院に入ったとき、市原先生に「これからはアミノ酸代謝では生きていけんよ。君はタンパク代謝にしなさい」と言われました。それでタンパク代謝を生体に近いシステムでやりたいということで、培養肝細胞の研究をやっていたんです。
先生もご存知のように酵素学としてのプロテアーゼ(タンパク質分解酵素)の研究というのは歴史は古かったんですけど、それではバイオロジーにはあまり結びつかない。私の最大の興味は、タンパク質にはそれぞれ寿命があるということで、タンパク代謝を研究するならこれが重要じゃないかと思っていたんです。そうこうするうち、77年にハーバード大学のまだ若い教授だったゴールドバーグが網状赤血球のタンパク質分解にエネルギーが要るという論文を発表しまして、これこそバイオロジーに結びつく研究じゃないかと。それで81年に彼のところに留学するんですけど、そのときはすでにゴールドバーグの系を用いてイスラエルのハーシュコたちがユビキチンのシステムを見つけていました。確かにタンパク質が壊されるとき、ユビキチンが結合するのにエネルギーを使うんです。ただ、留学している2年間の間に、ユビキチン化したタンパク質の分解もまた完全なエネルギー依存性であることがわかりまして、83年にエネルギー依存性には2つの段階があることを論文で書きました。それがきっかけだったですね。
岸本 プロテアソームの研究は、アメリカに行ってから始まったんですか。
田中 そうですね。当時、細胞内の分解酵素というのはほとんどリソソームが中心だったんですけど、細胞質に非常に巨大な酵素があるという報告がいくつかあったんです。それでエネルギー依存性の分解には2つの段階があるという論文を発表したときにゴールドバーグと話したのも、この巨大な酵素が関係しているかもしれないということでした。彼はこの酵素の生理学をやり、私はこの酵素の分子的実体を突き止めるということになって、日本に帰ってきたのですね。この酵素は不安定で誰も精製できなかったんですが、グリセリンを加えると非常に安定化するのを見つけて…。
岸本 プロテアソームの実体を精製されたわけですね。プロテアソームという名前は先生が付けられたんですか。
田中 名前はゴールドバーグと相談して、分離した酵素が普通じゃ考えられない大きさだったので、プロテアーゼ機能を持ったソーム(顆粒)ということで、88年に彼と連名で論文を出しました。それが最初ですね。
岸本 それがタンパク質としては一番大きなものですか。
田中 酵素複合体としてはたぶん一番複雑で大きいんじゃないかと言われています。その頃、生化学会でプロテアソームのことを発表しますと、構造を決めないとプロテアーゼかどうかわからないよと言われ、京大の中西重忠先生を紹介してもらいました。中西先生の部屋に行くと「プロテアソームなんて知らないよ」(笑)と。そこで説明すると、えらい興味を持ってくれまして総力を挙げて応援してくれることになりました。ともかくその酵素のバイオロジーというのは後にして、なぜエネルギーが要るのか、それがわからないということで、中西先生の遺伝子工学の技術を借りて、その後十数年かけてだいたい100ぐらいのサブユニットがあるんですけど、その7、8割を我々のラボで見つけていったんです。阪大の蛋白研の月原冨武先生にも協力していただいて、10年ぐらいかかったんですけど、触媒部位の構造解析もできまして、そうすると新しいタイプのスレオニン・プロテアーゼでした。なぜエネルギーが要るかですが、実はタンパク質の分解は加水分解反応ですから熱力学的法則としてエネルギーは要らない。結局、ユビキチン化したタンパク質を捕まえた後、触媒部位に運ぶときにタンパク質が立体構造を持っていると中を通過できないので、エネルギーはアンフォールディング、すなわち立体構造を壊してヒモ状にするために使っていることがわかったんです。
岸本 プロテアソームの構造は筒みたいになっていて、その筒がたくさんのサブユニットからできているわけですね。その前後に入り口と出口があるわけですか。真ん中で壊すわけですか。
田中 それはいつも聞かれるんですけど、幸いだったのは最初の電子顕微鏡写真を発表するとドイツのマックスプランク研究所のBaumeisterさんがプロテアソームの構造解析を一緒にやりたいと、徳島にやってきまして、それ以来今でも共同研究をさせてもらっています。彼の電子顕微鏡解析で、壊されるタンパク質にプロテアソームの入り口に入らないようなものを付けると、その両端に止まっているのでどちらも入り口だとわかりました。プロテアソームは、不思議なことが多かったんですが、ただ、タンパク化学の専門的な人たちからは興味を持っていただいたけれど、一般には変な酵素を研究しているなという程度だったですね(笑)。
岸本 2004年にユビキチンがノーベル賞をもらうまではそうだったですね。そのユビキチンが付いたタンパク質を分解するのがプロテアソームだということで、先生も朝日賞をもらい、日本学士院賞もおもらいになりましたね。
サブユニットの変換が起きる免疫プロテアソーム
岸本 あれはいつ頃だったですかね。新聞社から、インターフェロンで誘導される免疫プロテアソームというのが見つかったそうですけど記事にする値打ちはありますか、と電話があったとき、「僕は知りませんよ。それは何ですか」と答えました(笑)。インターフェロンが作用したら、プロテアソームの構造が変わるというけど何が変わるわけですか。
田中 プロテアソームはタンパク質を捕まえる調節ユニットと分解する触媒ユニットからできていますけど、触媒ユニットはα、βリングがα、β、β、αの順で結合していて、αリングは普通は閉じて無闇にモノが入ってこないようにブロックしているんです。触媒部位はβリングの内側にあります。
岸本 βのところで壊すわけ?
田中 βリングはβ1からβ7までのサブユニットからできていて、そのβ1、β2、β5に触媒部位があって、それぞれカスパーゼやトリプシンやキモトリプシンといった分解活性を持っている。ところがβリングは7つのサブユニットからできているのに脊椎動物の遺伝子は10あることに我々は気づいたんですね。それでインターフェロンで処理すると、3つのサブユニット(β1、β2、β5)が消えて3つのサブユニット(β1i、β2i、β5i)が現れることを見つけました。その遺伝子をクローニング(単離)すると非常に相同性があって入れ替わっていることがわかったので、免疫プロテアソームと名付けて94年に発表したんです。免疫システムというのは本当によくできていて、触媒のサブユニットを入れ替えることによって、感染したウイルスに由来するペプチド(タンパク質の一部)などの内在性抗原を効率的に切り出すことができるようになるんですね。免疫プロテアソームの発見は、プロテアソームのバイオロジーのはしりで、あれで多くの免疫学の人たちからも興味を持っていただきました。相前後してユビキチンの研究が爆発的に発展しまして、いっきに生命科学の中枢に。
岸本 胸腺プロテアソームやハイブリッドプロテアソームというのもありますね。
田中 胸腺プロテアソームは、胸腺皮質でのTリンパ球のポジティブセレクションに必要なペプチドを切り出す酵素として2007年に発表しました。
岸本 それもβリングのサブユニットが違うわけですか。
田中 そうです。β5が胸腺の皮質ではβ5tという独自なものに変わっていて活性がまったく違うんです。これらに関係して昨年、すごく大きな発見がありました。日本には皮膚の疾患で中条・西村症候群という全身性炎症をきたす難病がありまして、その原因が分かったんです。家系調査をして長崎大学と徳島大学のグループが別々に免疫プロテアソームのサブユニットβ5iに変異があることを見つけました。それを調べると、その変異が起こるとインターロイキン6が過剰に生産されて、それが病気の原因であることがわかりました。
岸本 なんでそのサブユニットが変わったら…。
田中 なぜというのがなかなか説明できなかったんですが、一方で我々はプロテアソームがどのように形成されるかを研究していまして、それが幸いしてその変異があると形成異常が起きてプロテアソームの機能が落ちることがわかりました。そうすると異常なタンパク質が蓄積して、ストレスが引き起こされインターロイキン6がものすごく増える。まさか私の研究が先生の専門のインターロイキン6に結びつくとは思っていなかったんですけど、それがプロテアソームの遺伝子変異による最初の疾患でした。

田中啓二 氏
東京都医学総合研究所所長
1949年、徳島県生まれ。72年徳島大学医学部卒業後、同大学大学院を経て76年同大学酵素研究施設(87年酵素科学研究センターに改組)助手。81~83年米国ハーバード大学医学部生理学部門に留学。95年徳島大学酵素科学研究センター助教授。96年東京都臨床医学総合研究所分子腫瘍学研究部門部長、2002年副所長、06年所長代行(先端研究センターセンター長兼任)。11年東京都医学総合研究所(臨床医学総合研究所、神経科学総合研究所、精神医学総合研究所が統合)所長。専門分野は、生化学、分子生物学。細胞内のタンパク質分解機構であるユビキチン・プロテアソーム系において、ユビキチン化したタンパク質を分解するプロテアソーム(プロテアーゼ複合体)の発見、構造・機能解析で世界的に知られる。受賞は、内藤賞、上原賞、朝日賞、東レ科学賞、武田医学賞、日本学士院賞、慶應医学賞ほか。
ユビキチンリガーゼが壊すタンパク質を見分けている
岸本 ユビキチン・プロテアソーム系のタンパク質分解で一番重要なのは、何でもかんでも壊すんじゃなくて目印の付いたものだけ壊すということですね。その目印がユビキチンというタンパク質を付けることで、ノーベル賞ももらいました。
田中 一般にタンパク質の立体構造はなかなか壊れなくて唯一の弱点は酸性なんですね。だから、小腸や胃でタンパク質はヒモ状に変性される。リソソーム内も酸性です。それまでの分解酵素というのは物理的な環境で変性したものを切るという形だったんです。ところがユビキチンの発見で考え方が大きく変わったのは、最初にこのタンパク質は不要だという識別があってそれを焼却炉みたいなプロテアソームに運ぶという、それが非常に新しかったですね。私はユビキチンの発見の現場にいたので、この言葉を最初に知った日本人です(笑)。当時、他の人はたぶん知らなかったと思います。
岸本 それで21世紀に入って壊すシステムが非常に重要やと。その1つはオートファジーですよね。オートファジーは何でもかんでもオートファゴソームに取り込んで壊す。先生のとは違う。違うけれども、最近はオートファジーも選択性があると言われていますね。それはどういうことですか。
田中 2つの分解システムには補完性がすごくあって、ユビキチン・プロテアソームの機能を落とすと、オートファジーの機能が上がるんです。なによりも細胞内にはミトコンドリアなどのオルガネラ(細胞小器官)がありますよね。これらの分解は全部オートファジーでして、異常なミトコンドリアを見つけて壊す。だから、基本的にオートファジーは非選択的ですけど、ある特定のものは選択的に取り込む。細菌なんかも侵入したら壊します。非常に不思議なのは受精すると、精子のミトコンドリアだけ消える。これもオートファジーなんです。
岸本 どうやって見分けるんですか。
田中 これらの場合、外膜タンパク質がユビキチン化されるんです。したがってユビキチンは、実はオートファジーのための分解シグナルとしても作用していることがここ2、3年の間にわかってきました。ただ、詳細についてはこれから研究が発展するんじゃないでしょうか。
岸本 それとね。遅かれ早かれ細胞内のタンパク質は壊されていくわけですけど、それぞれ寿命がありますよね。すぐに壊されるものと長い間、壊されないものと。その見分けはどうなっているんですか。
田中 それが私のライフワークなんですけど、結論から言いますと、タンパク質が不均一な寿命を持っている、その最終的なメカニズムはわかっていません。ただ、シグナル伝達や細胞周期のときにはリン酸化や酸化修飾が分解シグナルになるとか、いくつかのシグナルはわかっています。ゲノムの研究の恩恵でわかったのは、E3というユビキチンリガーゼ(ユビキチン連結酵素)の遺伝子がゲノム上に1000くらい存在することで、たぶんその酵素が相手を見分けていて、その見分け方として相手がリン酸化されていたりすると、素早くユビキチンがくっつくのですね。
岸本 どのタンパク質にはどのユビキチンリガーゼがというのも決まっているわけですか。
田中 もう100ぐらいは決まっています。実際、細胞周期やシグナル伝達に関しては、特有なE3酵素があるというのがわかっています。
岸本 そうすると、それがうまく働かないで病気になるというのは当然あっていいわけですよね。
田中 ユビキチンのシステムでの病気というのは、がんの場合とかたくさんあります。そういうのを“ユビキチン病”と呼ぼうというくらいあるんです。プロテアソームのほうは、変異すると早く死んでしまうのでなかったんですけど、免疫系のほうでは生存に必須じゃないので、中条・西村症候群のよう
な遺伝性疾患が出てきたんです。
岸本 病気はユビキチンリガーゼが異常を起こしてどれかのタンパク質が壊れなくなって起こるわけですか。
田中 それと、いったんユビキチンを付けたものを外す脱ユビキチン化酵素も沢山あって、こちらのほうの変異の病気がごまんと出てきていて複雑なんですね。
岸本 薬はどうですか。プロテアソームのほうの薬というと抗がん剤のベルケードですよね。これは効果があるんですか。
田中 ベルケードは今、血液内科でけっこう使われています。最初アメリカで使われ、現在、全世界で流行っています。プロテアソームの阻害剤ということで、今は武田薬品工業に買収されたミレニアム社に2回ほど呼ばれて行きました。どう思うかと聞くから「副作用が出るから絶対にダメだ」と言うと、帰ってくれと(笑)。10年ぐらい前にヨーロッパの学会で、ベルケードの投与でがんがこれだけなくなったとその開発者が発表したときも我々専門家は「あれはウソだよね」とか言っていたんです。ところが、ミエローマ(骨髄腫)は分泌性タンパク質が非常に多いので小胞体などに負担の大きい細胞なんですね。そんな細胞ではプロテアソームは30%ぐらい機能を落とすとアポトーシス(細胞死)のほうにシグナルが行くことがわかって…。
岸本 プロテアソームを働かないようにすると、異常なタンパク質が増えて細胞が生きられなくなるのを利用するわけですね。ミエローマ以外でも使えるということですね。
田中 ベルケードにはちょっと副作用があるので、現在、アメリカでは新しい阻害剤がいろいろ開発されています。今までの抗がん剤と作用機序が全然違うので、併用するといいんじゃないかという意見が多いですね。
岸本 先生はどこかで言っておられましたね。自分の研究は別に役に立つとか考えないでやってきたけど、こういう役に立つものも出てきたと。
田中 自分のやった研究が間接的に世の中の役に立つ。また、経済効果も生むというのはいいことだと思っています。研究を始めた当初は、そんなことはまったく考えもしなかったですけどね。
岸本 先生は徳島大学から東京に移られた。いつだったか、大阪大学に呼びましたけど、来られませんでしたね(笑)。
田中 結果的に私が東京に行ってよかったなと思うのは、やはり人材ですね。徳島にいても酵素学者としてプロテアソームはやっていたかもしれないけれど、今のように幅広くというのはできなかったでしょうね。
岸本 大阪ではあきませんか。
田中 いやいや(笑)。以前なら私は10なら10を全部知らないといけませんでしたけど、今は1ぐらいしか知らなくてあと9は弟子たちにまかせている。やはり私は人じゃないかと思いますね、研究というのは。だから、どうやっていい人を集めるか。
岸本 その集積効果が大事ですよね。
田中 今もいい人が集まる研究所にしたいと、そのためにできる限り研究しやすい研究所にしようとがんばっていますけど。
岸本 東京はもう何年になりますか。
田中 16、7年。ただ、なかなか言葉が江戸っ子にならなくて(笑)。最初に行ったときなんか、旧臨床研は東大の人ばっかりの研究所だったので、日本語でない言葉が流行っているぞと言われましたね。
岸本 そうでしたか。今日は、どうもありがとうございました。
EYES
ユビキチン化されたタンパク質を分解する プロテアソームを発見し、その構造・機能を解析
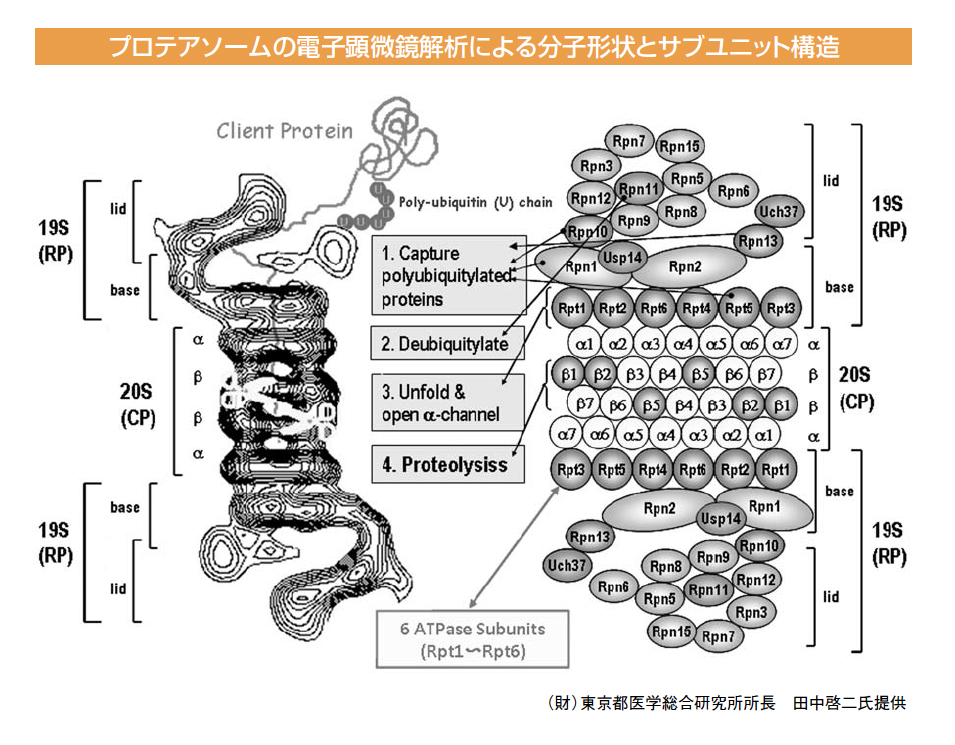
タンパク質の選択的分解機構であるユビキチン・プロテアソーム系における
プロテアソームの作動機構の解明を先導
細胞内では常時、必要なタンパク質が合成されるとともに、不要になったり、傷害を受けたタンパク質は分解されています。そうしたタンパク質の生と死のバランスの上に、増殖や分化、免疫反応など細胞の活動は成り立っています。遺伝子の情報に基づいてタンパク質がどのように合成されるかについては、遺伝子の本体であるDNAの発見以来、盛んに研究が繰り広げられ、その分子機構の解明も進みました。しかし、タンパク質の分解についてその機構解明の研究が飛躍的に発展し、大きな注目を集めるようになるのは、今世紀に入ってからでした。
その1つがオートファジーで、自食作用とも呼ばれます。細胞内にオートファゴソームという一過性の膜構造が現れて細胞質の成分を取り込み、分解酵素を蓄えたリソソーム(酵母や植物細胞では液胞)という小胞と融合して、取り込んだ成分を分解します。成分の中にはタンパク質だけでなく、ミトコンドリアなどの細胞内小器官も含まれています。オートファジーでは細胞質の成分が無差別に取り込まれるため、主にタンパク質などのリサイクルや細胞内の浄化に働いていると考えられています。
一方、不要になったタンパク質を選択的に分解するのがユビキチン・プロテアソーム系と呼ばれる分解機構です。ユビキチンという小さなタンパク質が標的となるタンパク質に数珠状に何個も付けられると、それを目印としてプロテアソームという酵素複合体が分解します。こちらはタンパク質専門で、この選択的な分解装置の発見が2004年のノーベル化学賞の対象となりました。このユビキチン・プロテアソーム系において、ユビキチンのパートナーとなるプロテアソームを発見し、その分子機構の解明に先駆的な役割を果たされたのが、今回、LF対談にご登場いただいた田中啓二氏(東京都医学総合研究所所長)です。
田中氏は、徳島大学酵素研究施設の助手だった1981年、米国ハーバード大学のゴールドバーグ教授のもとに留学し、真核細胞にユビキチン化されたタンパク質を分解するエネルギー(ATP)依存性の酵素が存在する可能性に着目。帰国後、その探索に取り組まれ、88年、ゴールドバーグ教授と連名でプロテアソーム発見の論文を発表されます。分子量250万という巨大な分子集合体でした(当時、最初に発見された触媒ユニットに対してプロテアソームと命名)。続いて、その構造・機能解析に何年にも渡って取り組まれることになります。田中氏が先導して明らかとなってきた全体像はおよそ以下の通りです。
プロテアソームは約100個のサブユニットから構成され、筒状の20S触媒ユニットの両端に19S調節ユニットが結合する形でユビキチン化されたタンパク質を分解します。調節ユニットはlid(蓋部)とbase(基底部)から成り、ここでユビキチン化されたタンパク質が捕捉され、脱ユビキチン化、アンフォールディング(タンパク質の立体構造の解きほぐし。エネルギーはここで使われます)されて、触媒ユニット内に送り込まれます。触媒ユニットはα、βリングがαββαの順でつながり、中は空洞になっています。通常αリングは閉じており、調節ユニットと結合することによって開きます。解きほぐされたタンパク質を分解する触媒活性はβリングの内側にあり、β1、β2、β5というサブユニットがそれぞれカスパーゼ、トリプシン、キモトリプシンという分解酵素のような活性を示します。
プロテアソームの構造・機能を解析する中で、田中氏はプロテアソームの分子多様性にも注目されます。インターフェロンの誘導によってβリングの触媒活性を担うサブユニットが入れ替わり、抗原提示に適応したプロテアソームを免疫プロテアソームと名付け、また最近、胸腺でのキラーT細胞のレパトア形成に関与する胸腺プロテアソームも発見されています。さらにプロテアソームを形成するための分子集合の機構解明も進められています。
田中氏は、ユビキチン・プロテアソーム系の一方の柱であるユビキチン、さらにオートファジーの研究にも取り組まれています。タンパク質の分解などに重要な役割を果たし、細胞の活動を支えるそれらの機構解明のさらなる進展が期待されます。