LF対談
シャペロンは小胞体に
タンパク質がたくさん来るときには増えないといけない No.71(2014.2)

京都大学大学院理学研究科 教授
森 和俊 氏
公益財団法人 千里ライフサイエンス振興財団
岸本忠三 理事長
分子生物学を学びたくてテキサス大学へ
岸本 僕が初めて森先生にお目にかかったのは、大阪科学賞の授賞式でした。その翌年にはガードナー賞を受賞される。僕自身は、ヒートショックプロテインという名前を知っているぐらいで、小胞体ストレス応答ってほとんど知らなかったんですけど、大阪科学賞のときに講演をされ、それがものすごい迫力でね(笑)。
森 そうですか。ありがとうございます。
岸本 アグレッシブな感じを受けました。そのときに印象に残っているのが、最初に京大の工学部に入って、それから薬学部へ。だけど、やっぱり理学部が世間からも注目されるとかで、「私は理学部の教授になりました」と言われていましたね。
森 そう言ったかどうか定かではありませんけど(笑)、もともと理学部に行きたかったんです。それこそ私が中学生の頃にクォークがたくさん見つかって、素粒子物理学がすごく華やかになりました。宇宙とか物質の根源に迫るような気がして、大学受験も京大理学部を目指していたんです。だけど、現役で合格する自信がなくて工学部へ。大学に入ってから、利根川進先生の分子生物学にすごく感銘を受けまして、生物学をやりたいと思うようになりました。
岸本 京大から岐阜薬科大の助手になられて、それからアメリカへ留学される。小胞体ストレス応答の研究は、アメリカに行ってから始まるわけですか。
森 そうです。
岸本 それまではまったく…。
森 タンパク質の高次構造形成っていうのが面白い問題だというのもまったく知らずに、「まあ、とにかくアメリカに行こう」ということでした。
岸本 じゃあ、なんでテキサス大学の小胞体ストレス応答をやっているところを選ばれたんですか。
森 私の先輩がテキサス大学に留学されていて、そこを紹介していただきました。テキサスならたぶん採ってくれるし、分子生物学を学ぶこともできると。とにかく分子生物学を学びたいという気持ちが強くて、テキサスへ。で、やってみたら非常に面白かったんで、そのまま4年半も居着いてしまったという感じです。
岸本 そのラボが小胞体ストレス応答の仕事を……。
森 ちょうど始めたところでした。
岸本 ヒートショックプロテインというのは?
森 歴史的にいうと、熱で誘導されるヒートショックプロテインがまずわかって、その研究から、細胞にはタンパク質の高次構造形成を助ける「シャペロン」というタンパク質が用意されていることがわかりました。
岸本 それはヒートショックプロテインだけではないわけですか。
森 小胞体の中のものは、よく似たファミリーですけど、熱で誘導されるわけではないです。それから、ホルモンとか受容体というタンパク質は細胞の表面に行く前に小胞体で形を整えるというのがわかってきたのも、1980年代の終わり頃でした。
岸本 小胞体で高次構造形成をする。
森 はい。で、私がアメリカに行ったのは1989年ですけど、その前年にテキサス大学の私のボスが、小胞体に形のおかしいタンパク質がたまると、核に情報が伝わってシャペロンが増えてくるのを見つけたところだったんです。
岸本 そういう概念はもう見つかっていたわけですか。
森 そうですね。
岸本 そこで、先生は形のおかしいタンパク質がたまったという情報を感知するセンサーを見つけられたわけですね、酵母を使って。
森 まず酵母のDNAにランダムに傷をつけて、異常なタンパク質がたまってもシャペロンを増やせないミュータント(変異体)を半年ぐらいかけて探して3種類見つけ、もう半年かけてそのミュータントを元に戻すような遺伝子を探すと、1つ見つかりました。それがセンサーでした。
岸本 それはどういう機構で、「変なものがたまっている」というのを見分けるんですか。
森 それはまだよくわかっていません。論文発表から20年以上経ちますけど。
岸本 その論文発表のときに、一緒にガードナー賞をもらわれた人との競争がありましたね。
森 ピーター・ウォルターですね。我々のその前に出した論文に目をつけて、突然入ってきたんです。
岸本 そっちのほうが後から。
森 後からですけど、先に93年6月号の『Cell』に論文を出されました。同じ酵母で、ほとんど同じやり方で。ただ、彼らはクローニング(単離)しただけで、リン酸化酵素や膜貫通型構造の証明もなかったので、異例だったんですけど、その2カ月後に我々の論文も『Cell』に載りました。で、その2つの論文でこの分野が始まったと言われていて、ワイリー賞とガードナー賞は彼と一緒にもらうことになりました。
ヒトでも小胞体ストレス応答は重要な役割を果たしている
岸本 我々の免疫の分野でいうと、抗体をたくさん作るBリンパ球のプラズマ細胞、哺乳類だったら、そういう細胞がいい研究のターゲットになるんですかね。
森 まさしくそうで、酵母で見つけた小胞体ストレス応答が、ヒトとか哺乳類でもちゃんと機能しているのであれば、きっとプラズマ細胞なんかで重要な役割をしているだろうと思っていました。
岸本 たくさん抗体のタンパク質を作るからと。
森 はい。センサーを見つけた後、シャペロンの発現を誘導する転写因子を酵母で明らかにして、その哺乳類バージョンも我々は見つけることができました。2001年に『Cell』に小胞体ストレス応答で活性化する転写因子だと報告したんですけど、それはハーバード大学のグリムヒャ―が見つけていたXBP1というものでした。
岸本 彼女のこと、よく知っていますよ、同じ免疫学で。
森 免疫系だけでそれをノックアウトしたマウスでは、プラズマ細胞に分化できないと彼女が半年前に報告していました。それは、我々からいうと小胞体ストレス応答がプラズマ細胞の分化に効いているということでした。
岸本 その転写因子がないと、プラズマ細胞にはなれないわけですね。哺乳類で実際にそういうふうに働いているのがわかったのは、プラズマ細胞だけですか。
森 酵母ではセンサーも1つ、転写因子も1つなんですけど、哺乳類になるとセンサーが3つになるんです。我々が酵母で見つけたIRE1というのは、酵母からヒトまで共通で、よく似ているものがあります。2つ目がアメリカのグループが見つけたPERKというもので、小胞体の中のタンパク質の状態がよくないときに、タンパク質を作りつづけるとますます状況は悪化するので、一時的に作るのを止めています。PERKをノックアウトすると糖尿病になります。
岸本 というと、インスリンを作る細胞で働いているわけですか。
森 はい、膵臓のβ細胞です。β細胞では、品質が悪くなるとPERKがブレーキをかけてタンパク質の翻訳を止めて調節しています。だけど、そのブレーキがなくなると悪いまま作りつづけるのでアポトーシス(細胞死)を起こしてしまうんです。それでインスリンが作れなくなります。
岸本 もう1つは何ですか。
森 ATF6です。これは我々が見つけたもので、センサー兼転写因子なんですね。ストレスを感知すると、転写因子ドメインが切り離されて核に行くという非常に面白いもので、そのノックアウトマウスを作ろうとやってみたところ、α、βと2つのフォームがあるんですけど、それをダブルノックアウトすると、非常に早く死んでしまいました。
岸本 いろんなところで働くということですね。
森 いろんなところで働きますし、3つのセンサーの中で発生の一番早い段階から重要です。なおかつ面白いのは、酵母ではIRE1の経路がシャペロンを誘導していますが、哺乳類になるとシャペロンを誘導するのはATF6のほうになります。IRE1-XBP1の経路は異常なタンパク質を分解処理するほうに働くようです。ただ、あまりにも早く死ぬので、もうちょっと見やすいメダカで解析して、昨年6年がかりで論文を出しました。背骨ができる前、体を軸として支える脊索が伸びるときにシャペロンが必要なんですね。非常に早い段階で影響が出るという論文を出したところです。
岸本 それはもうヒトにまで共通であるわけですね。
森 そう思っています。

森 和俊 氏
京都大学大学院理学研究科 教授
1958年、岡山県生まれ。81年京都大学薬学部卒業後、同大学院薬学研究科を経て、85年岐阜薬科大学助手。89年~93年米国テキサス大学に留学。93年(株)エイチ・エス・ピー研究所副主任研究員、96年主任研究員。99年京都大学大学院生命科学研究科助教授、2003年同大学院理学研究科教授。専門分野は生化学、分子生物学。細胞の分泌系タンパク質の品質管理を担う小胞体ストレス応答において、センサー分子や転写因子を発見、その分子機構や機能の解明を進める。受賞は、ワイリー賞、大阪科学賞、ガードナー国際賞、紫綬褒章、上原賞、朝日賞ほか。
ヒトの病気との関係を明らかにしたい
岸本 それほど生命にとってシャペロンというのは大事な仕組みなんですね。
森 そうです。面白いのは、シャペロンは1つひとつのタンパク質の形を整えるのに割と時間をかけてやっていますので、小胞体にタンパク質がたくさん来るときにはシャペロンの量も増えないといけないことです。
岸本 形を整えるというのはどういうふうに?タンパク質というのは、自然に立体構造をとるわけですよね。
森 それ、アンフィンゼンのドグマ(定説)といって自然にそうなるんですけど、たとえばタンパク質を構成する疎水性のアミノ酸間で不適切な相互作用が起きないように、そこをシャペロンがつかんだり、離したりしています。
岸本 おかしなところが引っつかないように…。
森 しているうちに…。
岸本 正常な形になる。
森 はい。つかんで離して、まだ疎水性の部分が露出しているとつかみ直して、疎水性の部分がきれいに構造の中に入ってしまうまで、これを繰り返すという感じです。また、あるシャペロンではカゴのようなものに入れたりします。だから、時間がかかるんです。
岸本 そうすると、さっきのプラズマ細胞ががん化したミエローマ(骨髄腫)では、シャペロンを増やせないようにするとか、分解にも回せないようにしたら、その治療に結びつくかもしれませんね。
森 抗がん剤ができないか。もうそういう取り組みはされています。海外でIRE1の活性を抑える薬の報告があって、実際、試験管内では効くんですけど、マウスにすると途端にダメになるらしいです。我々のところも同じ結果で、国内の中堅の製薬メーカーと協力して一生懸命スクリーニング(選別)してやったんですけど、細胞のレベルでは効いても、マウスにすると効かない。今、メダカの系ができましたので、最初から個体レベルでやってそこで効けば、マウスでも効くんじゃないかと考えているところです。
岸本 これから先は、どういう方向が目標になるわけですか。
森 やっぱり病気との因果関係をはっきりさせよう思っています。今世紀になって、いろんな病気に関与していると言われていましたので。今のところ、マーカー(指標)を測って、小胞体ストレスがかかっているかどうか、わかるまでなんですけど。
岸本 マーカーというのは?
森 小胞体のシャペロンのレベルを測るとかですね。3つのセンサーはどの組織にもあるんですけど、組織によって重要度が違います。膵臓のβ細胞にはATF6もIRE1もありますが、PERKがないとアポトーシスを起こします。肝臓だとIRE1のシステムが一番重要みたいです。
岸本 それぞれのセンサーが、どのタンパク質の異常を感知するのか…。
森 わかっていないです。それがわからないと、病気との関係はわかりません。
岸本 それをわかるには、どうしたらいいですか。
森 地道に仕上げていくしかないですね。今までは、小胞体ストレス応答の研究って薬を使ってやることが多くて、そうするとどのセンサーも活性化してしまいます。やっぱり生理的な状況というのは、どれか一番有効なセンサーが使われていると思われますので、それをきちんと調べていきます。
岸本 それが、これからの方向性ですか。
森 それと、その先に病気との関係を明らかにしていくということですね。
続けることを支援する研究費のシステムを
岸本 センサーが小胞体に異常なタンパク質がたまっているのをどうやって感知しているか、まだわかっていないということでしたけど、それはどうですか。
森 本当に何をもって異常としているかっていうのも、よくわからないんですね。また、90%以上のタンパク質はシャペロンに助けられて正常な形になるんですけど、10%以下のものはならなくて、それは小胞体から引っぱり出されて分解されます。そこもよくわかっていません。最初はなるべくシャペロンを使って直そうとするけれど、どうしても直せないと、時間が経ったものはもう壊していくようにしているのかもしれません。
岸本 センサーっていうのは受容体ですよね。それはどんな構造をしているんですか。
森 そのへんになると、パワーポイントでお見せしないと(笑)。
岸本 膜貫通型の内側の部分で認識しているわけですよね。何かのファミリーに属するとか、そういうことはないんですか。
森 小胞体タンパク質は、とにかくX線の結晶構造解析が非常に困難で、センサーに関しては解析できた例が2つしかないんです。1つは、ピーター・ウォルターが酵母のIRE1で解析したもので、二量体になっているところに溝があって。
岸本 そこに引っつく。
森 はい。そこに異常なタンパク質がはまり込むことによって活性化していると言っています。だけど、別の研究者はヒトのIRE1を解析して、そんなに溝は大きくないと。ここにははまらないと言っています。まだ、これからなんです。
岸本 そうですか。ところで、先生はこの分野では先頭を切っておられるわけだけど、研究費をとるのが大変やと言われていますね。そんなに難しいですか。
森 新学術領域というのに対応できなくて。我々はその前の特定領域研究というので3期サポートしていただきましたけど、今度のは「新」というぐらいだから、新しい学問領域じゃないとダメだと。
岸本 まだまだ新しい学問領域ですよね。
森 なんかもう一段落ついたように思われてしまっているんですよ。
岸本 ここからが始まりみたいなものですよね。
森 そうです。
岸本 続けてというのがね。外国だったらいい仕事をしとったら、ずっと続いていく。続けないといかんわけでね。次々と新しいこととか言わないで。
森 本当に特定領域から新学術領域への変化というのは、日本にとって決定的で、みんな言っています。人を育てるシステムになっていないと。若い人をインキュベートできないと。今の新学術領域では5年間はやれても、次の5年間は同じ延長上ではダメと言われるんですよ、たぶん。
岸本 続けていくことが大事やと思うんですけどね。そこから、どんどん発展させていく。僕の時代でも、1回だけだと言われて、「そこから続けることが大事なんや」と言って、続けてもらうことができた。
森 小胞体ストレス応答の研究を日本でやっていこうと思ったら、どうしてもグループの研究は通さないといけない。今度、私しか継続している者はいないグループを作って出し直してみたんですけど、それがどう評価されるか。それがダメなら、もう小胞体ストレスの灯が消えるかもしれないと思っています。
岸本 今日は、どうもありがとうございました。
EYES
タンパク質の品質を管理する小胞体ストレス応答という新分野を開拓
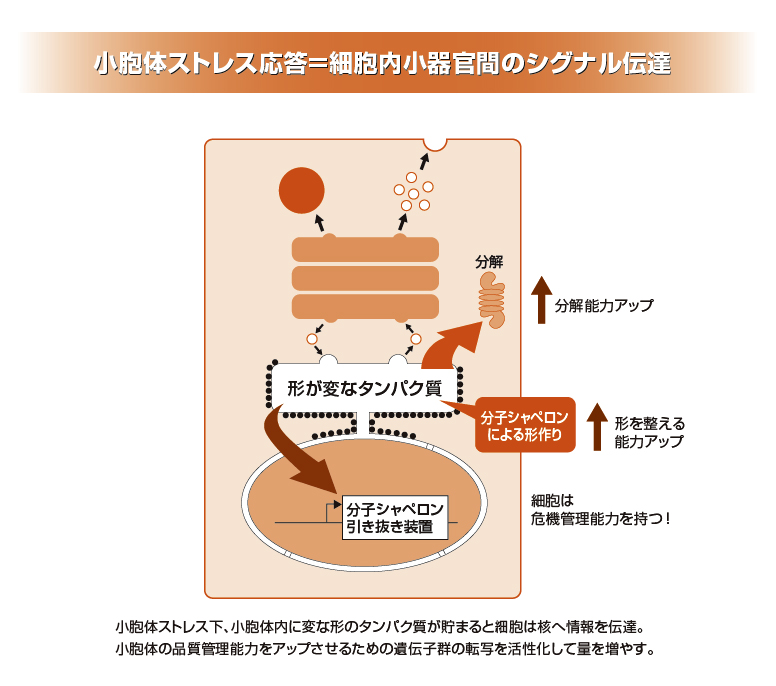
小胞体ストレス応答を支えるセンサー分子、転写因子を発見。
その精妙な仕組みの解明に挑む
酵素や膜受容体など、タンパク質はそれぞれ異なった正しい形(立体構造)を持つことによって生体内で特定の働きをしています。その形は、タンパク質を構成する各種アミノ酸の配列によって決まり、細胞で作られた当初はヒモ状だったタンパク質は自然に折り畳まれ、正しい形になると考えられていました。それは、アンフィンゼンの定説と呼ばれていました。
しかし、ヒートショックプロテインと呼ばれた熱によって増えるタンパク質の研究などから、1980年代後半にはタンパク質が正しい形になるのを助ける介添えタンパク質の存在が明らかになりました(ヒートショックプロテインは熱によって崩れたタンパク質の形を元に戻すために増えたのです)。1988年にはそうしたタンパク質は「シャペロン」と統一して呼ばれるようになります。シャペロンとは、若い女性が社交界デビューをするときに付き添う婦人という意味です。
一方、細胞にはミトコンドリアなどさまざまな細胞小器官(オルガネラ)が存在しますが、ホルモンなどの分泌タンパク質や膜受容体は、小胞体という細胞小器官でシャペロンによって正しい形に整えられ、細胞膜に運ばれることがわかったのも1980年代後半でした。そして88年には、小胞体の中に正しい形になれないタンパク質がたまると、シャペロンの量を増やすように核にシグナルを伝える「小胞体ストレス応答」という仕組みを細胞は備えていることが報告されます。その報告を行ったテキサス大学のゲシング教授のもとに89年に留学されたことをきっかけとして、小胞体ストレス応答を支えるセンサー分子や転写因子を発見するなど、その分野の開拓に多大な貢献をされたのが、今回、LF対談にご登場いただいた森和俊氏(京都大学大学院理学研究科教授)です。
森氏は、テキサス大学で酵母の小胞体ストレス応答の研究を進め、まずIRE1というセンサー分子を発見されます。IRE1は小胞体の膜を貫通する膜タンパク質でした。このセンサー分子が小胞体に正しい形になれないタンパク質がたまったことを感知するのです。その発見は、カリフォルニア大学サンフランシスコ校のピーター・ウォルター教授に先を越されましたが、93年、2カ月遅れて同じ科学誌『Cell』に発表されました。異例ですが、森氏の論文のほうが内容が詳細だったからです。この2人の論文によって小胞体ストレス応答の分野が実質的にスタートしたと考えられており、2009年、2人はガードナー賞を共に受賞されました。
森氏は帰国後も酵母で研究を進め、核のDNAにシャペロンを作ることを働きかける転写因子HAC1を発見されます。HAC1は活性化されたIRE1によって、そのmRNAがスプライシングされ、タンパク質として作られます。ここでも、ウォルター教授との激しい競争が繰り広げられました。続いて、森氏は酵母からもっと複雑な哺乳類に対象を移され、HAC1の哺乳類バージョンである転写因子XBP1、センサー兼転写因子であるATF6を発見されました。ATF6は小胞体の膜タンパク質ですが、活性化されると細胞質側が切れて転写因子として働くという、酵母にはまったくない仕組みを哺乳類は持っていることがわかりました。さらに、哺乳類ではPERKという膜タンパク質がセンサー分子として働き、これ以上小胞体に正しい形になれないタンパク質が増えないようにタンパク質全般の合成を抑制していることを海外のグループが突き止めています。
酵母ではIRE1-HAC1経路しかなかった小胞体ストレス応答が、現在、哺乳類ではIRE1-XBP1経路、ATF6経路、PERK経路と3種類あることがわかってきました。これらの経路はどのような役割分担をしているのか。また、小胞体ストレス応答と病気との関連はどうなのか。森氏もATF6の遺伝子を壊したマウスやメダカを解析することなどによって、それらの解明を試みられています。
小胞体ストレス応答では、タンパク質が正しい形になるようにシャペロンを増やしますが、どうしても正しい形になれないタンパク質は小胞体から引き抜き、分解されるようにするとも考えられています。ただ、その仕組みはまだよくわかっていません。また、センサー分子がどのように正しい形になれないタンパク質がたまったことを感知するのか、そのスタートのところもわかっていません。小胞体ストレス応答はまだまだ新しい分野です。今後の研究の発展が期待されます。