LF対談
学習判断と本能判断のあいだでの葛藤が、
人間の悩みの原点だと思います No.83(2018.2)

東京大学名誉教授、福井大学医学部特命教授
坂野 仁 氏
公益財団法人 千里ライフサイエンス振興財団
岸本忠三 理事長
免疫系の研究から神経系の研究へ
岸本 坂野先生は、京都大学でRNAasePの研究をされ、1976年にカリフォルニア大学サンディエゴ校へ移られました。そのときサンディエゴを訪れていた利根川進先生に会い、利根川先生の所属先だったバーゼル免疫学研究所に移られたんですよね。
免疫学の研究をすることになったのはどうしてですか。
坂野 そのころ、DNAの塩基配列決定やクローニングなどができるようになっていて、時代が大きく変わると感じ、高等動物の複雑系を研究したくなりました。ただ、神経系の研究が進むのはまだ先という感じがして、まずは免疫系から入ろうと思ったんです。
岸本 当時のご研究のハイライトは……。
坂野 『ネイチャー』のアーティクル4報に載った「免疫グロブリン遺伝子の体細胞組換え」の研究ですね。抗体遺伝子がどのようなルールで動き、多様性ができるのかを明らかにしたものです。
岸本 1982年にはカリフォルニア大学バークレー校に移られて、まだ免疫ですよね。
坂野 ええ。免疫系の研究では抗体遺伝子のシステムとしてのおもしろさは感じていたのですが、組み換え機構に関しては大筋で解明できて、「一段落ついたな」と思ったのです。
そのころから私は、ヒトの情動や行動の出力のしくみなどを分子レベルで明らかにしたいと考えていました。ちょうど、基礎生物学研究所を併任していまして、神経系の研究の立ち上げはそこで始めました。
鼻腔の受容体と嗅球の糸球体がつながり
「におい地図」が描かれる
岸本 身のまわりには、無数の種類のにおい物質があるわけですよね。受容体の遺伝子配列が変化して再構成するしくみがあると研究者たちは考えてきたわけですね。
坂野 ええ。マウスの嗅覚受容体は約1000種類。ヒトだと385種類しかありません。限られた種類の受容体で、種類が無限にあるにおいの情報をいかに識別するかということです。
岸本 どうやって識別するんですか。
坂野 嗅神経細胞は約300万個ありますが、1個の細胞は1000種類ある嗅覚受容体のうち1種類のみを発現します。そして嗅神経細胞から軸索が伸びて、大脳の前方下部にある嗅球表層に分布する1000個の糸球体の1つにたどりつきます。これにより、ある嗅覚受容体がにおいの情報を受けると、対応する糸球体が豆電球みたいに発火するんです。たとえば「1丁目1番地」の豆電球が発火すれば、1000種類のうちの「1種類目」の受容体が反応したからであり、「1丁目2番地」の豆電球が発火すれば「2種類目」の受容体が反応したから、という関係になります。
約1000個ある豆電球は、全体として見れば1000画素からなる電光掲示板みたいなものです。この電光掲示板は、嗅覚受容体が受けるにおいの種類によって、二次元のさまざまなパターンを描き出すことになります。「におい地図」とも呼ばれています。そのパターンを脳が認識することで、さまざまなにおいを識別しているわけです。
岸本 2004年のノーベル生理学・医学賞では、嗅覚の研究でリチャード・アクセルたちが受賞しましたね。でも、当時まだ、嗅覚受容体がにおい分子を受けてから、その情報がどこにどうつながるのか、詳しくはわかっていなかったんですよね。
坂野 そうです。1000種類の嗅覚受容体それぞれを発現している嗅神経細胞は、軸索を伸ばして1000の番地がある嗅球につながるわけですが、では軸索がどうやって1000ある糸球体のうちの1つを見分けるのかは未解明でした。
この問題に対する多くの見方は、糸球体側と嗅神経細胞側の両方になにか相互識別のしくみがあるのだろうというものでした。アクセルたちも当初、におい分子のようなものが糸球体の1000の番地それぞれに埋め込まれていて、伸びてきた嗅神経細胞の軸索の先端にも嗅覚受容体が発現していて、それにより番地が見分けられると考えていたようです。
けれども、それは実験的には支持されませんでした。
軸索の投射位置は相対的に決まる
岸本 坂野先生がそのしくみを解明したわけですが、結論はどのようなものでしたか。
坂野 たとえるなら、劇場のチケットに書かれている席番号を見て座席にたどりつくようなしくみではなかったということです。糸球位置は相対的なもので、体重も身長もちがう1000人がいて「体重もしくは身長の順に並べ」と言われて序列が決まっていくようなもの。この結果、糸球地図のトポグラフィーが決まるのです。
岸本 もうすこし詳しくお聞きすると……。
坂野 嗅球表面に形成される糸球地図には縦軸と横軸とがあります。縦軸とは背腹軸、横軸とは前後軸、つまり頭のほうを前、尾のほうを後とする軸です。
それで背腹軸については、その嗅神経細胞が嗅上皮の背側にあるか腹側にあるかで軸索が嗅球の背側にたどりつくか腹側にたどりつくかが、「ニューロピリン2」という軸索誘導分子を介して、決まることがわかったんです。
岸本 嗅神経細胞の位置と糸球体の位置が対応しているわけですね。では前後軸についてはどうなのでしょう。
坂野 実は前後軸のほうが難問で、背腹軸とちがって位置情報で決まるのではなく、発現している嗅覚受容体の種類で決まるんです。1000種類ある受容体が、一体何を手だてに軸索の投射位置を指令的に制御出来るのか、それが大きな謎でした。
岸本 そうなんですか。
坂野 嗅覚受容体などGタンパク共役受容体(GPCR)は、リガンド存在下では分子構造が変化してGタンパク質を結合し、環状アデノシン一リン酸(cAMP)を産生します。ところがGPCRはリガンドがない状態でも、Gタンパク質を結合することがあれば低レベルのcAMPを産生します。
この低レベルcAMPこそが、前後軸における投射位置を規定しているのです。ではこのcAMPの量はどう決まるかというと、嗅覚受容体の種類に固有な分子ゆらぎによるものである事が判りました。
岸本 嗅覚受容体の種類によって、cAMPの量が決まるわけですか。でも、どのように変換されるんですか。
坂野 GPCRはGタンパク質の結合部位が開いた活性型と、閉じた非活性型の2つの構造をとる事が知られています。リガンドのない状態でGPCRはこの2つの構造の間を行き来しているのです。我々は、熱運動による分子ゆらぎの間隙をぬって、Gタンパク質が入り込み、ノイズ程度のcAMPができると考えました。そのレベルは受容体の種類毎に異なり、それが軸索誘導分子「ニューロピリン1」の転写量を決めていたのです。
つまり、GPCRの分子ゆらぎによるノイズ活性が、嗅覚系では前後軸に沿った軸索投射を制御していたというわけです。

坂野 仁 氏
東京大学名誉教授、福井大学医学部特命教授
1947年生まれ。71年京都大学理学部卒業後、76年京都大学大学院理学研究科生物物理学専攻博士課程修了。76年京都大学化学研究所、日本学術振興会特別研究員。76年カリフォルニア大学サンディエゴ校化学部博士研究員。78年スイスバーゼル免疫学研究所研究員。82年カリフォルニア大学バークレー校微生物・免疫学部助教授。87年同准教授。87年理化学研究所主任研究員(併任1989年まで)。91年岡崎基礎生物学研究所教授(併任1997年まで)。92年カリフォルニア大学バークレー校分子細胞生物学部教授。94年東京大学大学院理学系研究科生物化学専攻教授。2012年東京大学名誉教授。13年福井大学医学部高次脳機能部門及び子どものこころの発達研究センター特命教授。嗅覚系における神経地図形成の基本原理の解明などで神経科学分野に大きく貢献してきた。専門分野は分子生物学、神経科学。趣味は登山。受賞はMERIT Award, NIH, USA、日産科学賞、東レ科学技術賞、武田医学賞、持田記念学術賞、内藤奨励賞、紫綬褒章。
「好き」「嫌い」は本能と学習の二重構造で決まる
岸本 おなじにおいでも、好きと感じる人もいれば嫌いと感じる人もいます。このことについても、坂野先生は研究を進めていらっしゃるんですよね。
坂野 ええ。好きと感じるか、嫌いと感じるか。感覚情報に対する価値決めのプロセスは、本能判断と学習判断の二重構造になっています。
本能判断とは淘汰の結果情報の価値決めが自然選択的に決まっている判断です。下等な動物では、感覚情報の価値判断はほぼ先天的に決まっています。たとえば、天敵臭や腐敗臭に対しては本能的に忌避します。高等動物と言われているヒトにも、本能的な価値判断は残っています。
一方、学習判断とは、記憶に基づいて入力情報の好き嫌いが判断されるというものです。
岸本 「いいにおいだけれど、たしか近づくと危険だったよな」とかですね。
坂野 ヒトの大脳皮質は肥大しているので、学習判断の占める割合は大きい。しかし、基本的な価値判断のしくみとしての本能判断もそのまま残されている。だから人間は、往々にして学習判断と本能判断のあいだで葛藤することになるわけです。これが人間の悩みの原点だと思います。世間体とかモラルとか宗教上の規範等も広い意味では学習判断に属しますから、本能的な欲望などに基づく先天的な判断とは相容れない場合が出てくるのです。私としては、この2つの対立する判断が、どこでどう裁定されるのかを神経回路レベルで明らかにしたい。
岸本 人間の心の問題とか、坂野先生の好きなところに入ってくるわけですね(笑)。本能判断と学習判断については、どう研究を進めたんですか。
坂野 まず、本能判断がどうやって決まるかを解明しようとしました。
2007年に行った「ネコを怖がらないネズミ」の研究では、マウスの糸球体地図の背側のほうだけを毒素を使って削ったのです。このマウスは残りの糸球体を使ってパターン認識による学習判断はできます。しかし天敵臭や腐敗臭に対して本能判断はできなくなってしまったんです。
この実験で私は「糸球地図というのは、におい情報をパターン化して識別するための画面であると同時に、においの質感ごとに領野分けもされている機能地図ではないか」と思ったのです。
最近我々は、キツネの匂い「トリメチルチアゾリン(TMT)」に反応する糸球体のうち、嗅球の背側奥にある1つを光遺伝学的手法を用いて発火させてみました。すると、そのマウスはフリージングという「すくみ反応」を示したのです。
TMT反応性の糸球体のうち、1個を光刺激するだけで恐怖行動が誘導されたわけです。いわばプッシュホンの短縮番号のようなものです。
岸本 本能的に「このにおいは好き」「嫌い」と判断がなされるとき、脳はどうなっているんですか。
坂野 嗅球を調べていくと、糸球地図の背側には、嫌いなにおいに反応する糸球が集まっていることがわかりました。これに対し腹側には、好ましく感じるにおいの情報が集められているのです。
これら、嗅球の背側に集められたにおいの情報は、扁桃体の皮質核につながっていて、そこを刺激するとストレス反応が誘導されます。一方、嗅球腹側に集められた情報は、扁桃体の内側核に送られ誘引的な社会行動を引き起こします。
つまり、本能的なにおいの好き嫌いは、入力した情報が一次投射によって仕分けられ、更に二次投射によって扁桃体の内側核にたどりつくか皮質核にたどりつくかによって、決まるという事がわかりました。
岸本 二次投射を決めているしくみのようなものはわかってきたんですか。
坂野 二次投射における軸索の配線は、一次投射の時と同じく、「ニューロピリン2」という軸索誘導分子が発現しているかどうかで仕分けられています。
このたった1種類の軸索誘導分子の発現で、においに対する本能的な好き嫌いが決まってしまうのです。
生後の短い時期における「刷り込み」が重要
岸本 坂野先生のお話を聞いて、嗅覚の研究は進んでいるんだなぁと感じました。
いまはどんな研究をされているんですか。
坂野 これはつい最近の研究ですが、生後まもない時期に嗅いだにおいというのは、たとえそれが忌避すべきにおいであっても、好きなにおいとして刷り込まれることがわかりました。
岸本 刷り込みですか。刷り込みがあると、嫌いなにおいに感じない……。
坂野 そうなのです。たとえば、生後まもないマウスに「4メチルティアゾール(4MT)」という、TMTに類似した本来は嫌がるはずのにおいを嗅がせてみます。すると生後1週間の臨界期にかぎって刷り込みが成立し、その後4MTに対して親近感を持つようになることがわかったんです。臨界期をたった1日外しただけで、刷り込み効果はあらわれません。
岸本 刷り込みの期間は短いんですね。
坂野 そうです。我々の最近の研究によって、刷り込みには一次神経が産生する「セマフォリン7A」が、神経活動依存的に関与する事が明らかになりました。一方、刷り込み記憶に対し誘引的な価値付けを行うのは、「オキシトシン」という脳内ホルモンであることも判明しました。
岸本 本能判断と学習判断が異なっているとき、脳はどうなっているんですか。
坂野 それが一番大事なところで、現在、扁桃体を中心に調べています。いまお話した、生後まもない時期に4MTを嗅いだ事のあるマウスでは、通常のマウス同様、4MTを嗅がせると本能回路を介した扁桃体皮質核の活性化が見られます。その一方で、普通のマウスであれば生じない扁桃体内側核の活性化が、刷り込み記憶によって生じていました。まさに、本能判断による「嫌い」と、刷り込み記憶による「好き」が扁桃体内でせめぎ合っているわけですが、この場合は「好き」のほうが勝ることになるわけです。
岸本生まれてまもないときの刷り込みの記憶は大事ということですね。
坂野 最近、幼児期の愛着保育が充分でない為に生じる愛着障害の問題がよく話題になりますね。私は、この精神発達障害にも刷り込み記憶が関わっているのではないかと見ています。事実、刷り込み記憶を成立させる「セマフォリン7A」のシグナルを臨界期に遮断したマウスでは、成長後に他の個体との関わりを避ける自閉症的な行動をとる事が示されました。
発育期に母親から語りかけられる、撫でてもらう、そして、においを嗅ぐ、そうしたことが、社会性のある健全な精神構造の発達に極めて重要ではないかということです。このあたりのことも今後研究していこうと考えています。
EYES
無数のにおい分子と1000種類の受容体が結合
二次元「地図」のパターンから 脳がにおいの質感を判断
嗅覚をめぐる2つの重要ルールを解明
好き・嫌い……感覚のしくみ究明にも挑む
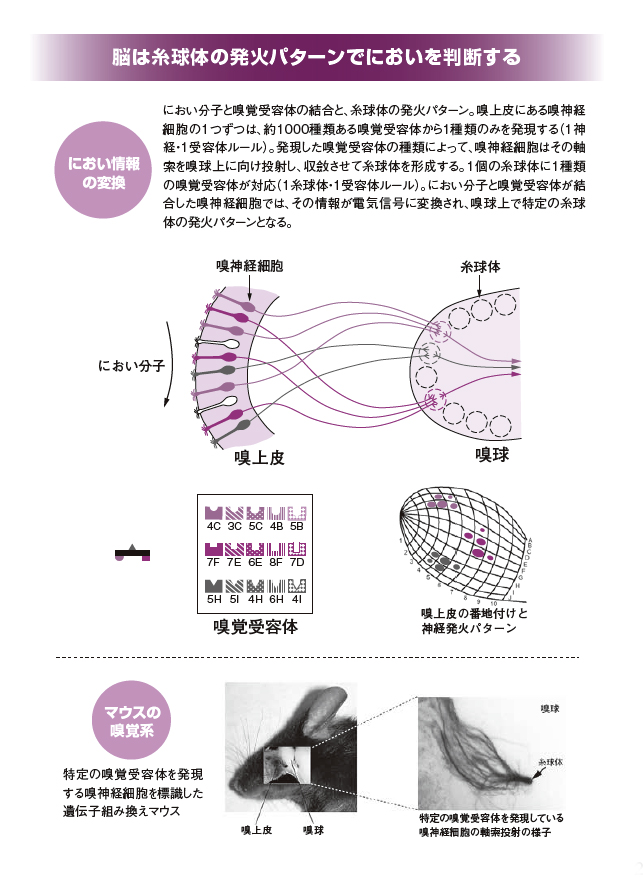
鼻に意識を集中させ、まわりの空気を嗅いでみると、どうでしょう。なにかしらの「におい」を感じるのではないでしょうか。私たちは日々さまざまなにおいに囲まれて生きています。においをもたらすのは、揮発性があり分子量300以下の低分子でできた「におい分子」です。数十万種類あるといわれています。
一方、におい分子を受けとめる生体の場所は、鼻腔の上部にある「嗅上皮」という粘膜状の組織です。嗅上皮には「嗅神経細胞」とよばれる細胞が集中しています。ヒトでは300万個ほど、マウスでは数百万個、また嗅覚の鋭さで知られるイヌでは数億個の嗅神経細胞があるといわれます。
嗅神経細胞には「嗅覚受容体」が発現します。嗅覚受容体は、特定のにおい分子と結合します。嗅神経細胞の数にくらべると、嗅覚受容体の種類はとても少なく、ヒトで約400種類、マウスでも約1000種類しかありません。しかし、1種類の嗅覚受容体が複数種類のにおい分子と結合できるため、数十万種類もあるにおい分子を相手にできるのです。
嗅神経細胞と嗅覚受容体の間には「1神経・1受容体」という重要なルールがあります。1個の嗅神経細胞に着目すると、1000種類ある嗅覚受容体のうちたった1種類しか発現していないのがわかります。もし、1個の嗅覚神経細胞が複数種類の嗅覚受容体を発現してしまったら、どの受容体にどういうにおい物質が結合したかがわからなくなるので、これを避けているわけです。
嗅覚受容体とにおい分子が結合すると、その刺激は電気信号となって嗅覚神経細胞から軸索を伝い、脳の前方にある「嗅球」に向かいます。そして、嗅球の表面には「糸球体」とよばれる球状の部位があり、その数はマウスで約1000個。さきほど、嗅覚受容体の種類は約1000と話しましたが、数が一致するのは偶然ではありません。1個の糸球体は1種類の嗅覚受容体と厳密に対応しているのです。こちらは「1糸球体・1受容体」ルールとよばれます。
1000個の糸球体の1個ずつは、対応する嗅覚受容体がにおい分子と結合した場合に発火します。たとえば、図において「4C」という嗅覚受容体ににおい分子が結合すると、嗅球上にある糸球体の「4-C」が発火するわけです。また「5H」の嗅覚受容体がにおい分子と結合した場合、糸球体の「5-H」が発火します。
対談記事では、糸球体を“電球”に、またそれが1000個ほど集まったものを“電光掲示板”にたとえています。この電光掲示板は、におい分子と嗅覚受容体の結合の組み合わせによりさまざまなパターンを画像として示すわけです。二次元平面上に表されたパターンは「におい地図」ともよばれます。
脳の中枢は、この電光掲示板に示されたパターンをその都度、捉えています。それにより、どういうにおいが入ってきたか、さらに、そのにおいは自分にとって良い(好きな)ものか悪い(嫌いな)ものかを判断しているのです。
今回の対談に登場する東京大学名誉教授の坂野仁氏は、「1神経・1受容体」と「1糸球体・1受容体」というルールが分子レベルでどう成立しているかや、軸索がにおい地図のどの位置に投射するかといった、嗅覚をめぐる重要なしくみをつぎつぎと解明してきました。坂野氏の研究は着実に進み、近年はにおいの情報から「好き」「嫌い」などの感覚が引き起こされるしくみの解明にも挑んでいます。どんな成果が上がっているか、対談を覗いてみましょう。